Biologie Moléculaire et Cellulaire
MORT CELLULAIRE : APOPTOSE ET NECROSE
La mort cellulaire a été mise en évidence tardivement. A l'heure actuelle, c'est un processus
obligatoire à toute cellules Eucaryotes et organismes pluricellulaires.
On est en train de mettre en évidences un processus semblable chez les Bactéries.
Cette M.C.P. (Mort Cellulaire Programmée) est responsable de l'hémostase cellulaire qui
apparaît dès le début de la vie d'un individu (cf. article).
Notons quelques pathologies de la régulation de cette M.C.P.: cancers (perte du verrou
apoptotique), maladie d'Alzheimer (atteinte u système cérébral), maladie de Thurtington,
S.I.D.A., ...
1. Importance de la M.C.P.
Définition de la M.C.P.
Processus « cellulaire actif » qui atteint son paroxysme lors de la M.C.P. Cette M.C.P. Peut se produire suite à des signaux issus du développement ou de l'environnement ou encore en réponses à des liaisons physiologiques décelées par le réseau de surveillance interne de l'organisme (système immunitaire).
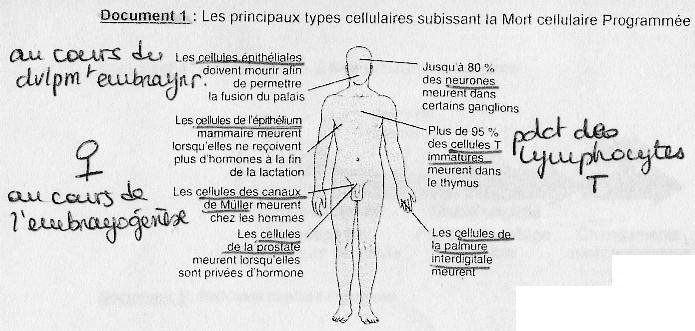
Principales cellules qui subissent la M.C.P.
cf. poly A doc.1Cellules nuisibles:
En priorité: les Lyphocytes T autoréactifs qui agissent contre le soi (si persistance = pathologie auto-immune), les cellules à ADN muté, les cellules qui abritent des agents pathogènes.
Cellules « inutiles » :
Par exemple les neuroblastes au cours de la vie embryonnaire (neurones en excès, précurseurs
des neurones). Seuls les neuroblastes ayant déjà établis des connexions nerveuses persistent.
Aussi: les cellules de la palmure interdigitale (aucune utilité chez l'Homme), les canaux de
Müller chez les hommes.
Lors des traitements de cancers par chimiothérapie, les molécules agissent en provoquant des
liaisons cellulaires qui vont déclencher la mort cellulaire par M.C.P.
M.C.P.: Apoptose – Nécrose.
Il existe deux voies principales par lesquelles les cellules meurent:
-> voie accidentelle = Nécrose,
-> voie programmée = suicide actif dont le principal mécanisme est l'Apoptose (cf. article
M.C.P. 1, 2, 3).
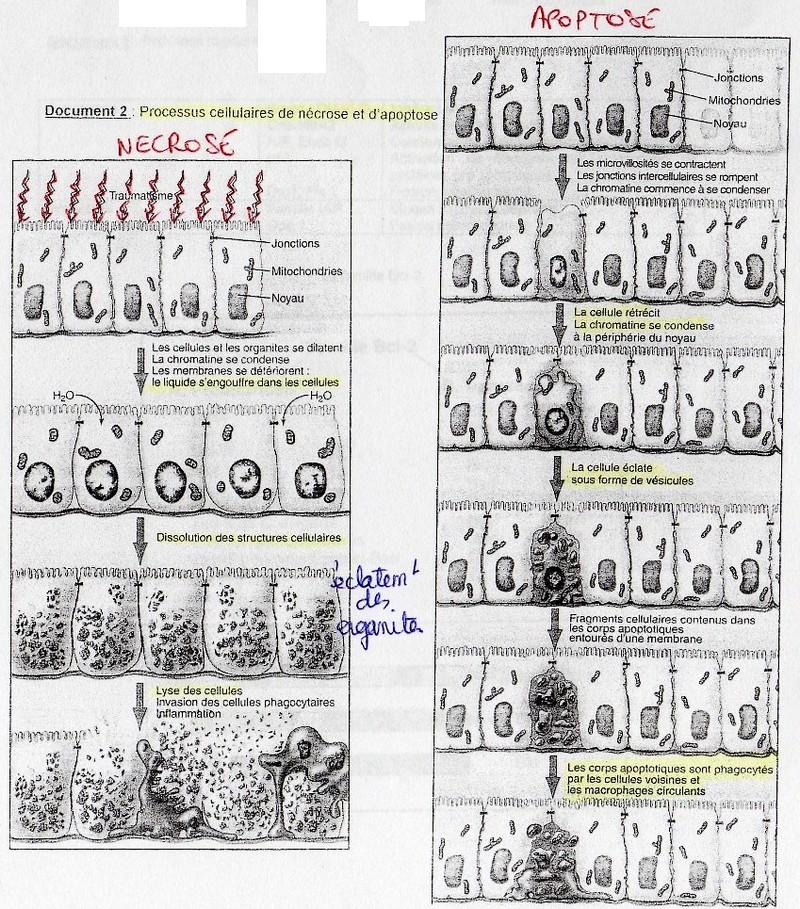
Nécrose:
cf. doc. 2 poly. ALa nécrose se produit quand la cellule reçoit, ou subit une agression structurale ou chimique
dont elle ne peut pas se remettre. Ex: ischémie (= manque d'oxygène ), température extrême,
traumatisme physique,...
Globalement la cellule se désagrège et est digérée par ces propres enzymes ( autodigestion). Il
y a alors une pénétration massive d'eau dans la cellule. Les cellules gonflent, les organites
vont éclater. Ce qui libère les enzymes lytiques, les lysosomes et les péroxysomes entraînant
une la digestion de la cellule et donc sa lyse. Il y a alors une forte inflammation locale; ses
molécules mobilisent les cellules phagocytaires qui vont nettoyer la zone nécrosée.
La nécrose s'étend à cause de la présence de phagoytes.
Apoptose:
Ici la cellule se condense sur elle-même. La chrnologie est très reproductible ici. Ce qui permet
d'établir que ce phénomène est régulé et qui provient de la cellule. (cf. doc. 2 poly. A)
Premier phénomène: modifications des jonctions cellulaires, elles se rompent (pour les tissus).
Alors la cellule se recroqueville, se condense sur elle-même. Ce qui entraîne une perte de
l'asymétrie des membranes. Le PhosphatydylSérine (marqueur membrane) se retrouve en
même quantité sur les deux feuillets alors que normalement il n'y en a que sur le feuillet
interne. La cellule éclate sous forme de vésicules composés de débris cellulaires appelés "corps
apoptotiques". Parallèlement on observe une hypercondensation de l'ADN.
Enfin, tous ces sorps apoptotiques sont phagocytés par deux types de cellules: les cellules
voisines de l'apoptose et les macrophages circulants.
Ici il n'y a pas de réaction inflammatoire car les débris sont contenus dans des vésicules,les
cellules voisines ne sont donc pas dégradées.
Comparaison Nérose – Apoptose:
cf. doc. 3 poly A2. Mécanisme moléculaire de l'apoptose.
M.C.P.: Apoptose – Nécrose.
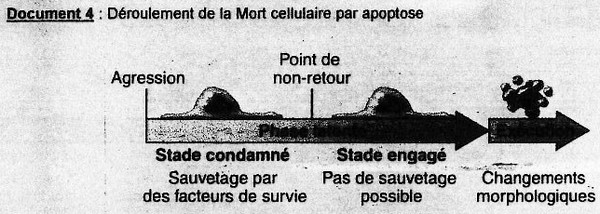
cf. doc. 4 poly. BL'apoptose débute toujours par un signal interne à la cellule (coupure dans l'ADN, stress
oxydatif, infection virale,...) Un signal externe peut-être à l'origine de l'apoptose (diminution
de facteurs de croissance, contact avec des molécules toxiques, rayonnements X,UV ou g. Ces
signaux sont dits "pro-apoptotiques".
[Mécanisme détaillé en suite.]
Phase de latence:
Durant cette phase, les cellules ont un aspect normal au niveau physiologique mais ont quand
même une activité pour les gènes impliqués dans l'apoptose. (Dt variable de quelques heures à
quelques jours).
Il y a un point de "non-retour". Auparavant = "stade condamné", où la cellule peut encore
arréter l'apoptose s'il y a beacuoup de facteurs de survie. Ensuite = "stade engagé", où la
cellule va être détruite obligatoirement.
Cette première phase est régulée par de nombreux protéines notamment p53, molécule qui
contrpole l'état de l'ADN, puissant activateur de l'apoptose, molécule dite "supresseur de
tumeurs" ou "anticoncogène".
Phase d'exécution:
Ici se s'effectuent beaucoup de changements physiologiques et morphologiques (phase très rapide » 1 heure).
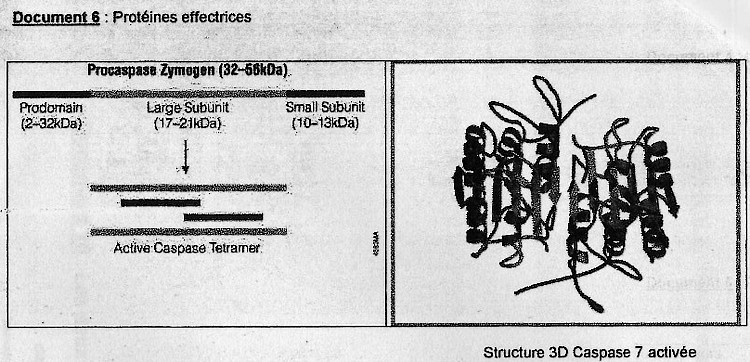
Protéines impliquées dans l'apoptose.
De nombreuses protéines sont impliquées dans ce mécanisme. On distingue:
Les protéines régulatrices:
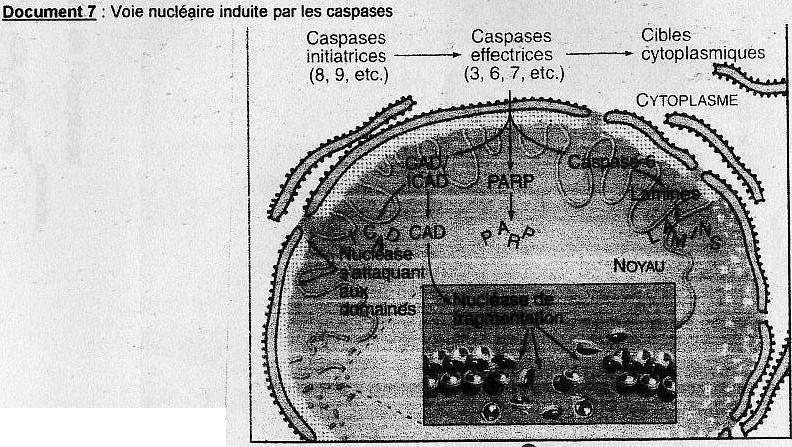
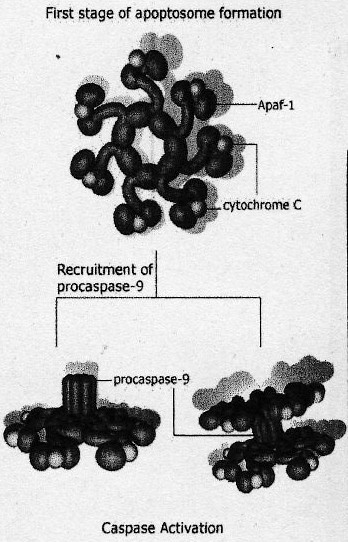
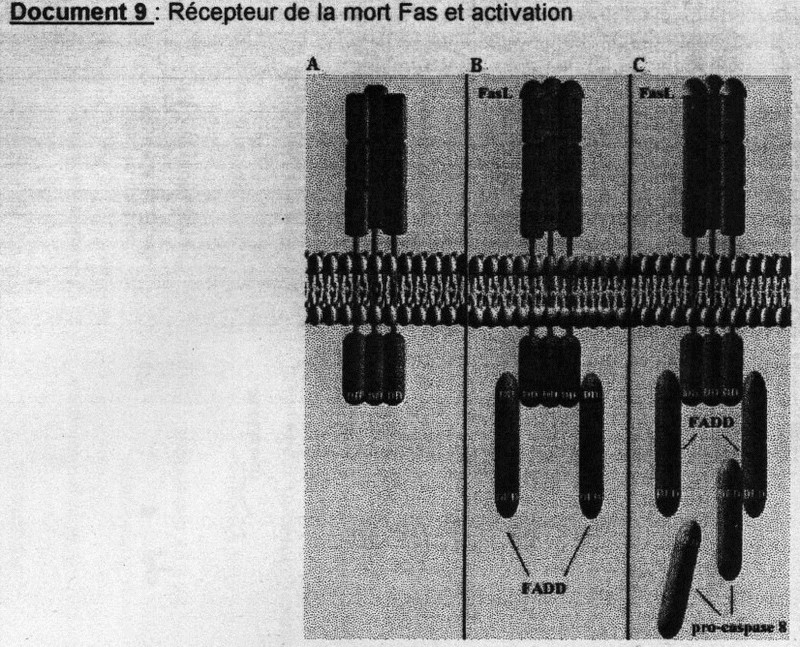
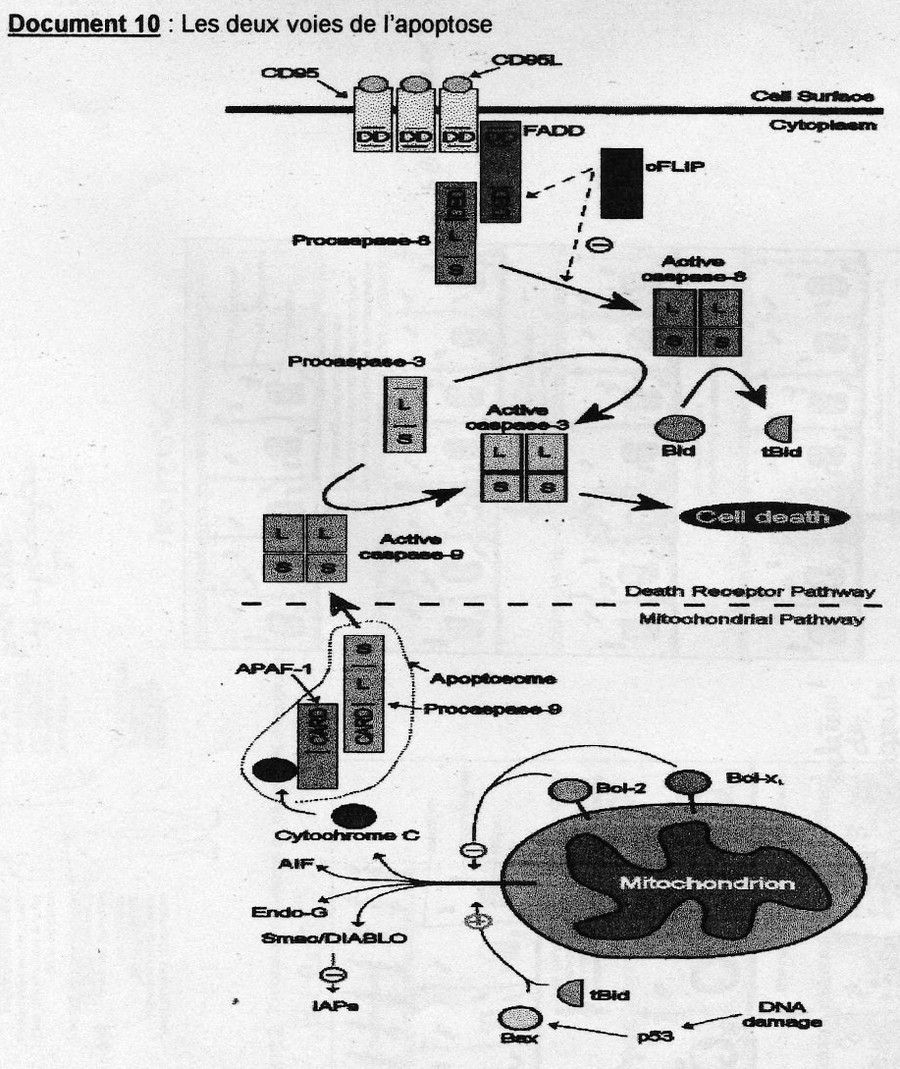
(cf. doc. 5 tiré de l'article)On retrouve des protéines activatrices ou pro-apoptotiques et des protéines inhibitrices ou aniapoptotiques. Parmis cette grande diversité de protéines une famille y tient un rôle central: les protéine sde la famille Bcl-2 (possèdent toutes des domaines d'homologies BH pour Bcl- Homology très conservé). Ex: Bax et Bak, favorisent le relarguage d'effecteurs contenus dans l'espace intermembranaire de la mitochondrie soit en formant un pore membranaire ou en activant sa formation. Bcl-2 semble bloquer la relarguage du cytochrme C par la mithochondire.
Cycle cellulaire du MPF:
Le MPF induit la phosphorilation de nombreux systèmes celulaires: comme l'histone H1
(nucléosome) qui, en se phosphorilant, s'associe à la fibre nucléosomale et favorise le
compactage de l'ADN; comme les laminas nucléaires sur la face interne de l'enveloppe
nucléaire; comme les Topoisomérases II qui sont des enzymes du processus de réplication,
elles déroulent l'ADN.
La tansistion G2 – M n'est possible que si:
la réplication de l'ADN est totale, si il y a une bonne réplication alors il y a liaison sur l'ADN.
SPF.
Il est le contrôleur du passage "Start" constitué d'une cycline de phase G1 et d'un Cdk. Il active les gènes des protéines impliquées dans la réplication. Il régule l'adénine des ADNpolymérases et va contrôler la bonne duplication des centromères.
3. Régulation du Cycle Cellulaire.
Elle implique des facteurs extracelulaires qui sont les "facteurs de croissance" (hormones, protéines qui stimulent la croissance cellulaire). cf. SVF
Définition.
Facteurs de croissance:
Les facteurs de croissance sont, par défintion, une susbstance protidique qui agit sur un événement du cycle cellulaire. Généralement entraîne un effet stimulant. Sont des molécules soit produites par les cellules environnementales, soit autoproduites.
Différentes catégories:
cf. tableau poly. ELes facteurs de croissance peuvent avoir un effet "ubiquitaires":
PDGF -> dérivés des plaquettes,
EGF -> dérivés des tissus épidermiques,
Erythropoietin -> hormone qui stimule la production des globules rouges.
Au niveau de leur mode d'action, on distingue 3 types de facteurs de croissance:
- • facteurs de croissance de "Compétence",
• facteurs de croissance de "Progession",
• facteurs de croissance de "Survie".
Les facteurs de Progression sont indispensables sur le point de restriction ("start"). Ils n'agissent que si la cellule a été préalablement sensibilisé par les facteurs de Compétences. Les facteurs de Compétences permettent de donner des conditions favorables aux futur cycle cellulaire. Ces facteurs de Compétences stimulent l'expression de récepteurs spécifiques aux facteurs de Progressions.
Donc les facteurs de Compétences et de Progression stimulent le cycle cellulaire.
Les facteurs de Survie agissent après le cycle cellulaire et inhibent l'apoptose (à G0). Ex: Fibroblastes: 3T^3 fibroblastes de souris repiqués au 1/3 tous les trois jours. Au bout de 3 semaines on observe une crise apoptotique = 99% de cellules mortes et 1% de survivants.
Ces dernières sont des cellules dites transformées qui sont devenues insensibles aux facteurs apoptotiques (lignées établies = ensemble de cellules génétiquement identiques et définies). En culture cellulaire ces trois types de croissances sont nécéssaires (-> SVF).
Transduction des signaux de cellules.
cf. poly. EFacteurs qui agissent via un récepteur membranaire:
Il existe trois récepteurs membranaires différents:
• récepteurs à Tyrosine Kinase qui s'autoactive une fois ligand fixé sur un récepteur.
• récepteurs liés aux protéines G, ces facteurs produisent un message seondaire (AMPc,
IP3, DAG).
Tous ces signaux issus des récepteurs membranaires induisent des enzymes cytoplasmiques:
"sitch kinases" qui stimulent, en phophorilisant des acteurs du cycle cellulaire.
SRF+CREB: facteurs cytoplasmiques activateurs de gènes régulateurs du cycle cellulaire.
Facteurs qui possèdent un récepteur intracellulaire:
Exemple des hormones lipophiles qui traversent la membrane mais qui doivent avoir un
récepteur cytoplasmique (pour les hormones stéroïdiennes) ou nucléaire (pour les hormones
thyroïdiennes). Ces récepteurs agissent sur le génome directement.
En culture cellulaire ces facteurs de la croissance sont apportés par le SVF qui va permettre, in
vitro, de donner les conditions favorables à la prolifération cellulaire: toutes privations de SVF,
in vitro, va bloquer la division cellulaire et inexorablement déclencher l'apoptose.
La division cellulaire est parfaitement contrôlée et régulée afin que ce processus se déroule le
plus parfaitement possible. Toute varation de ces régulations induit la transformation cellulaire
dont le premier état est l'immortalité, c'est-à-dire la division infinie.
4. Annexes.
Schémas