Biologie Moléculaire et Cellulaire
CYCLE CELLULAIRE : CONTRÔLE ET REGULATION
Prérequis: Phénomène de Mitose acquise.
Les cellules se reproduisent en duplicant leurs constituants et en se divisant par deux. On distingue deux cas:
➢ Organismes cellulaires et procaryotes: ici on arrive à deux individus identiques; on a n division qui donnent 2n individus.
➢ Organismes pluricellulaires: ici il y constitution d'un individu « mature »= adulte c'est-à dire capable d'être autonome pour son fonctionnement biologique et capable de se
reproduire. Au sein de cet individu mature, les divisions cellulaires vont être régulées avec pour but:
-> de renouveler les cellules à durée de vie limité;
-> de maintenir à peu près constant le nombre de cellules;
-> d'empêcher les proliférations anarchiques (tumeurs par ex.).
Tout ceci constitue l'Hémostasie cellulaire.
Caractéristiques de la division cellulaire:
➢ obtenir une cellule fille identique, c'est-à-dire un ADN parfaitement répliqué et des chromosomes parfaitement partagés entre les deux cellules;
➢ la masse cellulaire et des organites doit être doublée;
➢ tout le processus doit être régulé et surveiller pour éviter toute erreur.
1. Grandes Phases du Cycle Cellulaire.
Présentation.
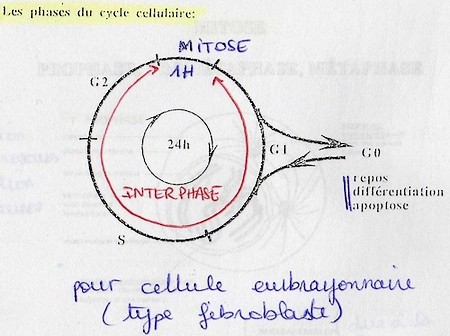
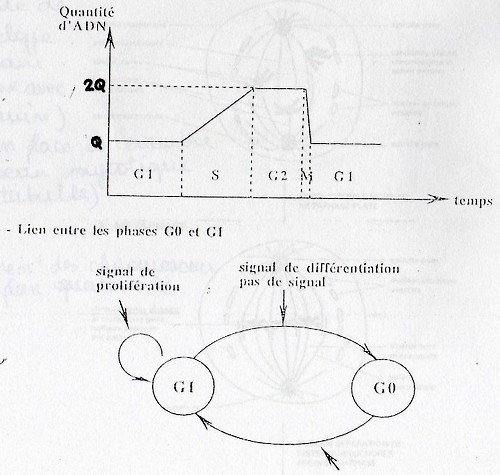
Le cycle cellulaire est composé de différentes phases qui se succèdent dans le temps: G1 – S – G2 – M. Elles peuvent alterner avec une phase particulière: G0 directement connectée à G1.
En G0 la cellule est au repos, différenciée ou la cellule déclenche l'apoptose; la cellule n'est pas en division.
Durée du cycle:
G1 – S – G2 sont des phases de préparation à la mitose, elles ont des durées variables.
G1 et G2 font parties de l'interphase.
La durée du cycle cellulaire dépend de la nature de la cellule et aussi de son âge:
→ cellules épithéliales: renouvellement de la peau, tps = 10h pour le cycle;
→ cellules hépatiques: tps = 1 an pour ce répliquer;
→ cellules type neurones: celules extrêùeùent différenciées donc pas derenouvellement (max de neurones à 15-18 ans).
Les cellules les plus rapides sont cellules de l'embryon, tps = 1h (car ici que de la mitose).
Processus nucléaire:
cf. diagramme La mitose divise le doublement de l'information génétique dans les deux cellules filles.
Différentes phases.
Phases G1 et G2:
"G" = Gap = liaisons qui servent à prépare les étapes suivantes.
➢ G1 prépare la synthèse de l'ADN. Ici, la cellule reste sensible aux facteurs environnementaux (notamment aux facteurs de croissance). La celule poursuit son
cycle cellulaire de façon autonome au-delà de G1.
➢ La transition de G1 à S est provoquée par l'augmentation de la taille de la cellule jusqu'à une taille critique.
➢ G2 prépare la mitose, c'est-à-dire qu'il s'agit de l'élaboration de tous les constituants
qui participent à la mitose (ici, duplication des centrioles).
Phases S:
"S" = Synthèse. C'est ici que le patrimoine génétique de la cellule est dupliqué.
Phases M:
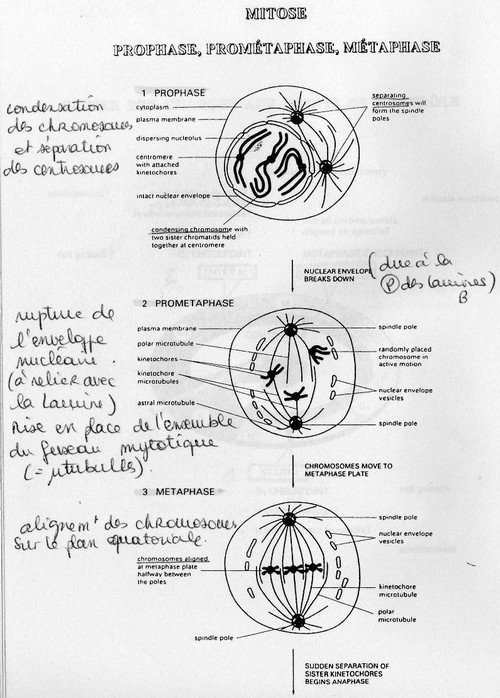
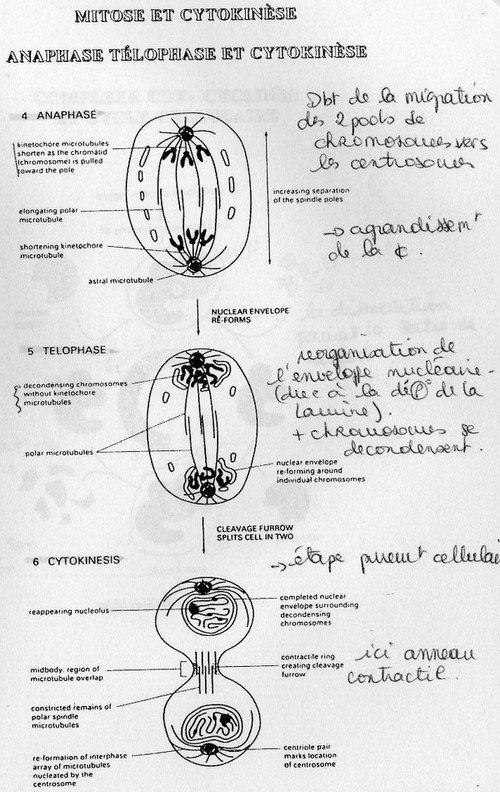
Phase de division à proprement parlé = Mitose.
Elle est divisée en cinq étapes nucléaires et une étape cellulaire. La métaphase est la clef de la
régulation. L'étape cellulaire est la "cytodierèse" où la cellule mère se divise en deux cellules filles.
[connaître le processus de la Mitose, cf. doc. B]
Processus Cellulaire.
Duplication des centrosomes:
Elle se déroule avant l'étape de Mitose. Cette duplication est fondamentale pour la génèse du fuseau mytotique. Ce processus débute en phase G1, les deux centrioles se séparent ce qui permet le début de la duplication. Cette duplication se poursuit pendant S et G2. Avant la fin de G2, on a deux centrosomes fonctionnels (=centre de polymérisation des microtubules mytotiques). Pendant la Mitose, les deux centrosomes migrent aux 2 pôles de la celule.
Duplication des organites:
Ce processus se déroule généralement avant la Mitose et au niveau cellulaire. Les organites présents se fractionnent de manière à obtenir 2 pools d'organites dans la cellule mère. Il peut y aoir synthèse de novo ( = élaboration de nouveaux organites pour mitochondries et cholroplastes). Pendant la phase de cytodiérèse, ces 2 pools d'organites seront répartis équitablement dans les deux cellules filles.
2. Contrôle du Cycle Cellulaire.
Il s'agit d'un mécanisme purement intracellulaire, indépendant des facteurs extracellulaires. Les mécanismes vont expliqués comment vont s'enchaîner les différentes phases du cycle.
Concept de Contrôleur.
Il existe deux théories qui s'opposent:
-> Chaques étapes du cite cellulaire est responsable du déclenchement de la suivante.
Hypothèse: à chaque étape il y a un contrôleur?
-> Modèle du "Supercontrôleur" c'est-à-dire un ensemble moléculaire qui supervise l'ensemble
du cycle. Il faut donc contrôler les étapes clefs: G1, M et la sortie de la Mitose.
En fait, le contrôle du cycle se fait par ces deux systèmes. C'est-à-dire que chaque étape
possède son contrôleur et l'ensemble des contrôleurs est supervisé par un supercontrôleur. Un
meilleur contrôle est ainsi assuré ce qui limite au maximum les erreurs éventuelles.
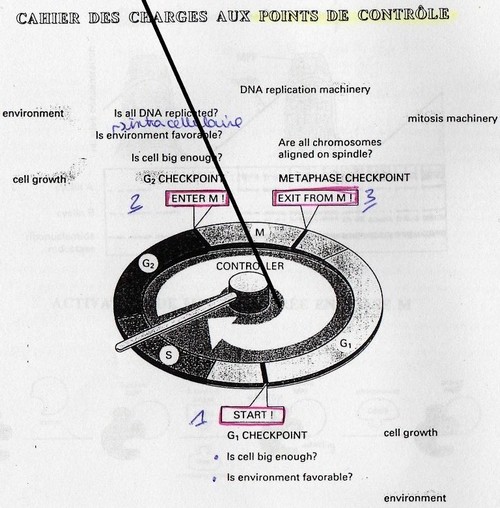
Les points de contrôles:
Il y a trois points de contrôles.
- 1er point: à la fin de G1, le point "Start" permet de contrôler deux évenements à savoir si
l'environnement est favorablement à la duplication et si la taille de la cellule est suffisante.
Pour la taille on regarde le rapport RNP. Tant que ces conditions ne sont pas favorables, la
cellule ne peut pas passer en division. Dès que ce point est franchit la cellule devient
indépendante de l'environnement ("pont de non-retour").
- 2ème point: à l'entrée de phase en M. Ici on vérifie que tout l'ADN a bien été répliqué et qu'il
y eu une bonne réplication. Cela est contrôlé par P53, Rb (protéines de contrôles) qui sont des
molécules anti-oncogènes c'est-à-dire supresseurs de tumeurs. On vérifie la quantité et la
qualité de l'ADN répliqué.
- 3ème point: lors de la Mitose, pendant la Métaphase on vérifie que les chromosomes sont
parfaitement alignés sur le plan équatorial.
Si la cellule note une anomalie à un point de contrôle alors il y a arrêt du cycle cellulaire et
déclenchement de l'apoptose.
Les contrôleurs:
De nos jours on a isolés des contrôleurs pour chaque phase et un supercontrôleur (MO15). Il a
été isolé par une équipe française à Montpellier.
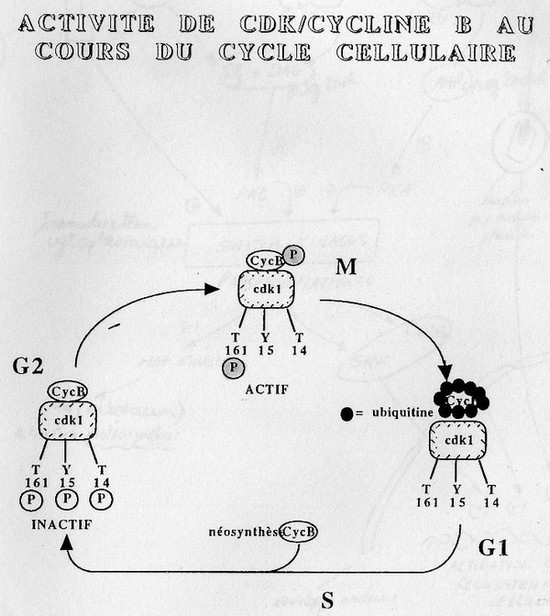
Chaque contrôleur de phase est un hétérodimère constitué d'un CdK (Cyclin Dependant
Kinase) et d'une partie régulée = la Cyclin, elle est la protéine spécifique. Cet hétérodimère
n'est fonctionnelle que lorsque le Cdk et la cyclin sont associées. Pour chaque phase il y a deux
principaux contrôleurs MPF (M phase Promoting Factor) pourla transition G2 -> M, SPF ( Start
Promoting Factor) pour le passage de G1 -> S.
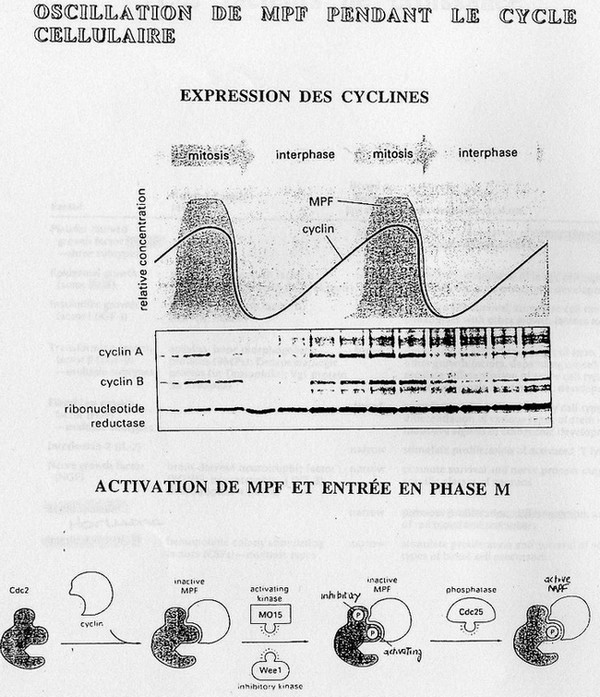
MPF.
Oscillateur pendant le cycle:
cf. poly B[Même phénomène pour SPF]
La ribonucléotide réductase est le témoin interne, il s'agit d'une protéine exprimée en quantité
constante quelque soit le cycle. Elle quantifie de façon relative la quantité en Ca et Cb avec
une densité des traits. Cette manipulation a été réalisé avec des cellules synchrones, c'est-àdire
qui ont la même période de cycle cellulaire (la synchronisation de fait par une brève
culture dans un milieu sans sérum).
Pour que le MPF soit fonctionnel il faut qu'il soit phosphorilé sur la cycline et qu'il y ait un
phosphate sur le Cdk. Pour le désactiver, il faut détruire la cycline associée. Cette
désactivation est induite par l'ubiquitine.
[ubiquitine: petite protéine cellulaire qui s'associe avec les protéines qui doit être détruites ( =
marqueur) et qui seront détruites par le protéasome].
Ces phosphorilations sont effectuées par Mo15, le Spercontrôleur.
Le MO15 permet de réguler le MPF par la phosphorilation variable.
Cycle cellulaire du MPF:
Le MPF induit la phosphorilation de nombreux systèmes celulaires: comme l'histone H1
(nucléosome) qui, en se phosphorilant, s'associe à la fibre nucléosomale et favorise le
compactage de l'ADN; comme les laminas nucléaires sur la face interne de l'enveloppe
nucléaire; comme les Topoisomérases II qui sont des enzymes du processus de réplication,
elles déroulent l'ADN.
La tansistion G2 – M n'est possible que si:
la réplication de l'ADN est totale, si il y a une bonne réplication alors il y a liaison sur l'ADN.
SPF.
Il est le contrôleur du passage "Start" constitué d'une cycline de phase G1 et d'un Cdk. Il active les gènes des protéines impliquées dans la réplication. Il régule l'adénine des ADNpolymérases et va contrôler la bonne duplication des centromères.
3. Régulation du Cycle Cellulaire.
Elle implique des facteurs extracelulaires qui sont les "facteurs de croissance" (hormones, protéines qui stimulent la croissance cellulaire). cf. SVF
Définition.
Facteurs de croissance:
Les facteurs de croissance sont, par défintion, une susbstance protidique qui agit sur un événement du cycle cellulaire. Généralement entraîne un effet stimulant. Sont des molécules soit produites par les cellules environnementales, soit autoproduites.
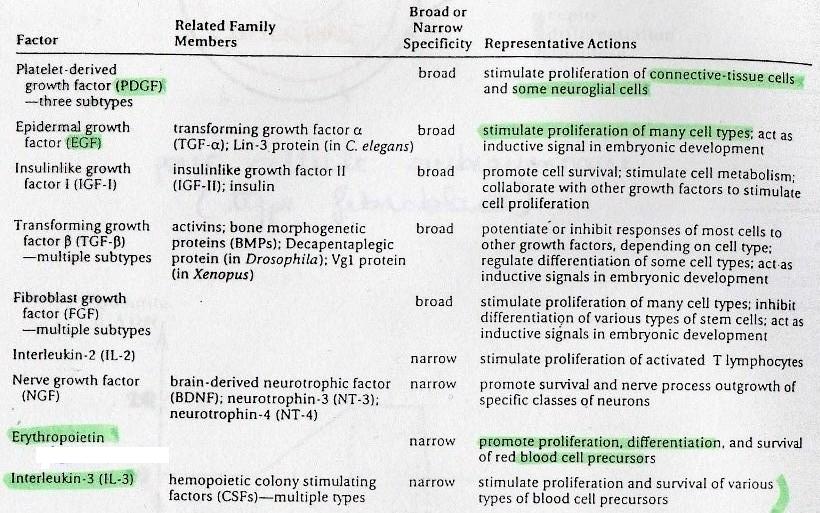
Différentes catégories:
cf. tableau poly. ELes facteurs de croissance peuvent avoir un effet "ubiquitaires":
PDGF -> dérivés des plaquettes,
EGF -> dérivés des tissus épidermiques,
Erythropoietin -> hormone qui stimule la production des globules rouges.
Au niveau de leur mode d'action, on distingue 3 types de facteurs de croissance:
- • facteurs de croissance de "Compétence",
• facteurs de croissance de "Progession",
• facteurs de croissance de "Survie".
Les facteurs de Progression sont indispensables sur le point de restriction ("start"). Ils n'agissent que si la cellule a été préalablement sensibilisé par les facteurs de Compétences. Les facteurs de Compétences permettent de donner des conditions favorables aux futur cycle cellulaire. Ces facteurs de Compétences stimulent l'expression de récepteurs spécifiques aux facteurs de Progressions.
Donc les facteurs de Compétences et de Progression stimulent le cycle cellulaire.
Les facteurs de Survie agissent après le cycle cellulaire et inhibent l'apoptose (à G0). Ex: Fibroblastes: 3T^3 fibroblastes de souris repiqués au 1/3 tous les trois jours. Au bout de 3 semaines on observe une crise apoptotique = 99% de cellules mortes et 1% de survivants.
Ces dernières sont des cellules dites transformées qui sont devenues insensibles aux facteurs apoptotiques (lignées établies = ensemble de cellules génétiquement identiques et définies). En culture cellulaire ces trois types de croissances sont nécéssaires (-> SVF).
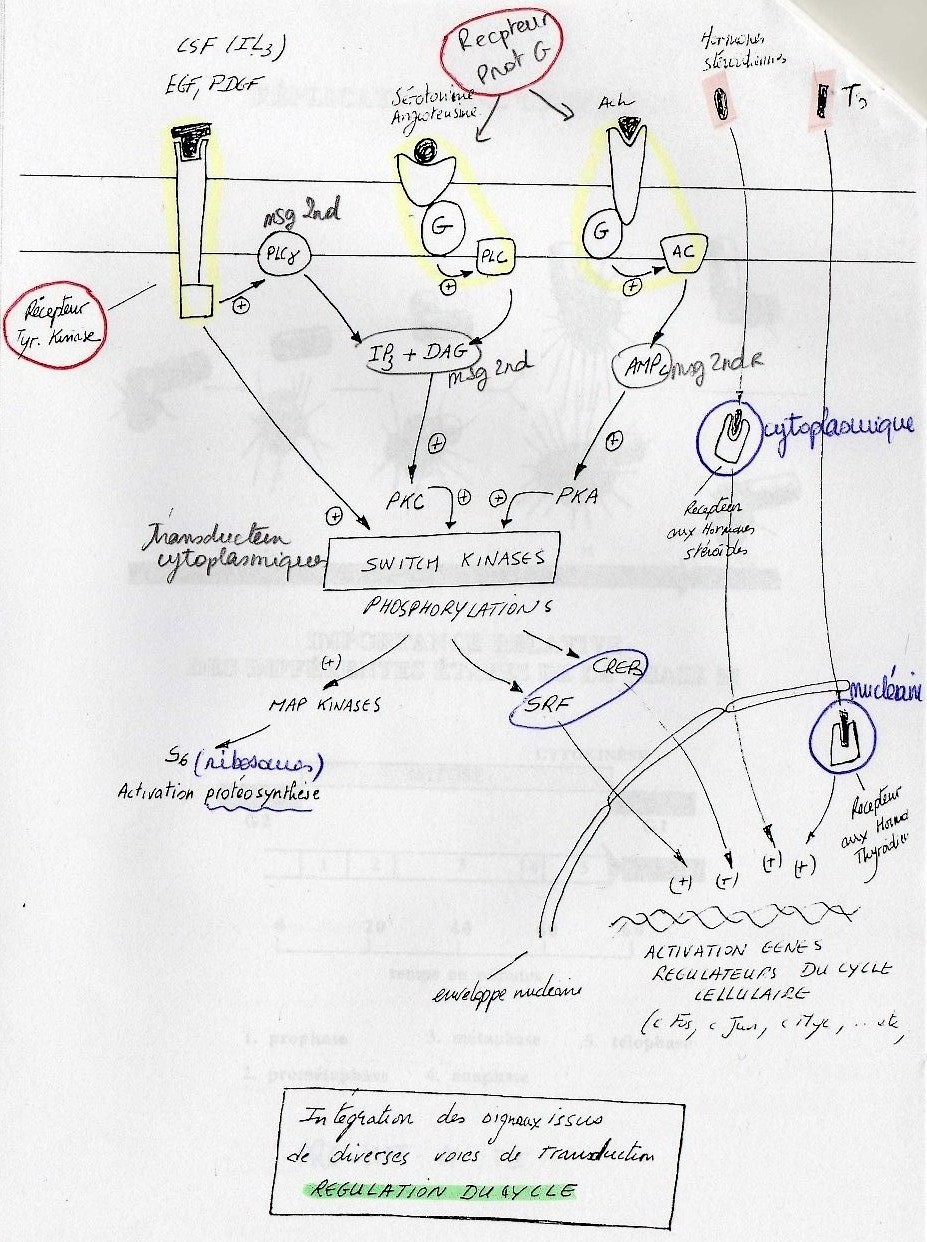
Transduction des signaux de cellules.
cf. poly. EFacteurs qui agissent via un récepteur membranaire:
Il existe trois récepteurs membranaires différents:
• récepteurs à Tyrosine Kinase qui s'autoactive une fois ligand fixé sur un récepteur.
• récepteurs liés aux protéines G, ces facteurs produisent un message seondaire (AMPc,
IP3, DAG).
Tous ces signaux issus des récepteurs membranaires induisent des enzymes cytoplasmiques:
"sitch kinases" qui stimulent, en phophorilisant des acteurs du cycle cellulaire.
SRF+CREB: facteurs cytoplasmiques activateurs de gènes régulateurs du cycle cellulaire.
Facteurs qui possèdent un récepteur intracellulaire:
Exemple des hormones lipophiles qui traversent la membrane mais qui doivent avoir un
récepteur cytoplasmique (pour les hormones stéroïdiennes) ou nucléaire (pour les hormones
thyroïdiennes). Ces récepteurs agissent sur le génome directement.
En culture cellulaire ces facteurs de la croissance sont apportés par le SVF qui va permettre, in
vitro, de donner les conditions favorables à la prolifération cellulaire: toutes privations de SVF,
in vitro, va bloquer la division cellulaire et inexorablement déclencher l'apoptose.
La division cellulaire est parfaitement contrôlée et régulée afin que ce processus se déroule le
plus parfaitement possible. Toute varation de ces régulations induit la transformation cellulaire
dont le premier état est l'immortalité, c'est-à-dire la division infinie.
4. Annexes.
Schémas