Biologie Moléculaire et Cellulaire
TRANSFERT DE PETITES MOLECULES AU TRAVERS DE LA MEMBRANE
La cellule Eucaryote a besoin d'eau, de sels minéraux et de macromolécules pour vivre
(production d'énergie et source de matières premiaires.). Ils pénètrent de façon différentes
dans la cellule, suivant sa structure ou particuarité physico-chimique: sa taille (150 kDa =
limite d'exclusion), caractères hydrophobes et hydrophiles, charge de la molécule.
La cellule a adapté à chaque élément une modalité d'échanges.
Rôle des transferts de molécules: approvisionnement en nutriements, gestion des déchets,
régulation des volumes cellulaires, maintient du pH, de la force ionique (fondamental pour la
survie de la cellule), maintient des gradient électro-chimique ( -70V à – 90V, potentiel de
repos).
La memebrane cytoplasmique est perméable de facon sélective, on dit que cette perméabilité
est "différentielle" (la vitesse varie selon la molécule), cette perméabilité est orientée.
1. Différents modes de transferts.
Les mouvements des molécules sont de 3 types:
Diffusion Simple (ou Libre).
Définition:
Mouvements libre de particules au travers de la membrane régit par la loi de la diffusion.
DIFFUSION: phénomène par lequelles moléules d'une phase de dispersion ( c'est-à-dire le
solvant) et d'une phase dispersée ( c'est-à dire le soluté) tendent à occuper le maximum de volume et à avoir une répartition homogène. Le mouvement se fait suivant le gradient de
concentration.
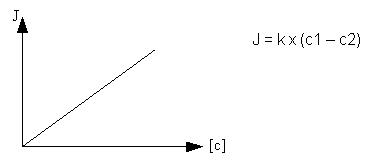
Loi de Fick:
J = dn/dt = - Ps x A x (c1-c2)
où
(c1-c2) = différence de concentration entre les deux compartiments définis par la membrane.
A = surface d'échange, zone d'échange où la molécule traverse la membrane.
Ps = perméabilité spécifique de la molécule considérée.
J = flux net, quantité quantité de matière qui passe au travers de la memrane par unité de temps et de surface.
Molécules impliquées dans ce mode de transfert:
La molécule impliquée doit avoir un poid moléculaire inférieur à 150kDa (cad 150 g/mol), cela correspond aux gaz respiratoires, l'eau, acides gras et composés comme le glycérol. Ce genre de molécule doit être hydrophobe.
Transport Facilité.
Mise en évidence:
On met E. Coli en présence de glycérol marqué au tritium H3 , on étudie alors la quantité au
sein des 2 compartiments.
Le tracé ressemble à un tracé enzymatique, ce qui ne correspond pas à une diffusion libre; on
peut donc conclure que E. Coli possède une protéine de transport.
J= ( Jmax x [S]) / ( KM + [S])
où S = glyérol.
Le mouvement se fait dans le sens du gradient de concentration (cad du plus concentré au
moins concentré). Le transporteur, à savoir une protéine, est plus ou moins spécifique de la
molécule transférée. Ce mode de transport est plus performent que la diffusion libre. Comme il
y a un transporteur c'est alors un système saturable (limitation de nombre de transporteur).
Ce mode de transfert est sensible aux facteurs physico-chimique car il ressemble à une
réaction enzymatique. Cependant il n'y pas de dépense d'énergie, et tout type de molécule
peut subir ce transport (sauf les gaz).
Transport Actif.
Définition:
Ce mode de transport nécessite un transporteur spécifique de la molécule véhiculée. Le
mouvement se fait dans le sens inverse du gradient de concentration, il nécessite donc une
dépense d'énergie.
Il existe le transport actif direct dit primaire et le transport actif indirect dit secondaire.
Ce mode de transport est sensible aux inhibiteurs respiratoires (NaN3, Cyanure CN, ...).
C'est également un système saturable.
Différents transports actifs:
(cf. Poly. A doc. 2)- Pompe: transport actif pimaire.
- Uni-Sym-Antipore: transport actif secondaire.
→ Uni: 1 molécule , sens unique
→ Sym: 2 molécules, dans le même sens
→ Anti: 2 molécules, sens opposés
2. Structure des transports, Rôles biologiques.
Pour les transports facilité et actif.
Trois types de structures.
Pompes:
Ce sont des protéines à activité enzymatique qui utilisent soit l'hydrolyse de l'ATP, soit
l'énergie lumineuse pour transférer les molécules. Leur fonctionnement met en place un gradient de concentration au travers de la membrane.
Ex: ATPase Na+/K+, Bactériorhodopsine, ATPase H+.
Elles présentent une spécificité absolue vis-à-vis de la molécule transportée.
Mouvement contre le gradient de concentration.
Transporteurs:
Ce sont des protéines qui fournissent des voies de transferts facilités selon les gradients de
concentrations. Le transferts se fait par modification conformationnelle de la protéines
transporteuse (cf. Doc. 1 modèle "ping-pong"). Il n'y pas de dépense d'énergie. Certains
transporteurs font partis intégrante du transport actif secondaire (ex: sympore, antipore).
Leur spécificité est variable; le mouvement se fait dans le sens du gradient, parfois le transfert
d'une autre molécule de fait dans le sens inverse.
Canaux:
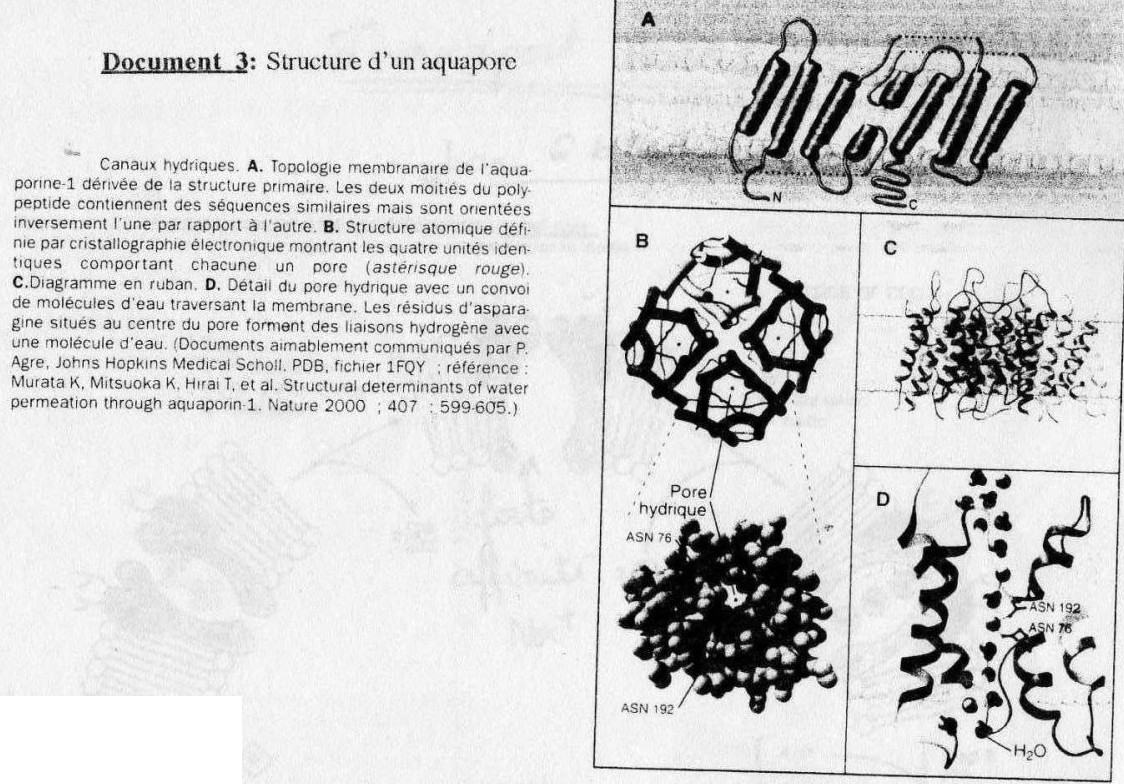
Ce sont des structures de transport. (cf. Doc. 3)
Ils présentent une forte spécificité vis-à-vis d'un ion. L'ouverture et la fermeture des canaux
est transitoire sous l'ation d'un mécanisme de régulation. Lorsque le canal est ouvert: flux
massif dans le sens des gradients électrochimiques.
Ex: porines chez les Procaryotes.
Quelques fonctions biologiques.
Phénomène d'osmose:
L'osmose a été étudié quaond on voulait savoir comment l'eau traversait la membrane cellulaire.
→ Ps H2O = Pcalculée (d'après la loi de Fick)
→ Pmesurée (vessies de batracies)
Pm >> Pc
Il existe au niveau des membranes des systèmes de transferts d'eau plus performants que la
diffusion libre.
On a traité à l'ADH (αDiurétique Hormone) les vessies de batraciens pour favoriser la
réabsorption de l'eau. On constate que Pm > Pc.
Le témoin sans ADH montre que Pm ≈ Pc.
Ce système de transfert d'eau est dépendant d'un signal chimique inducteur, ce sont les
aquapores (structures isolées du transport membranaire) = canaux de 0,7 n de diamètre qui
sous l'ation de signaux chimique tels que l'ADH sont capables de se regouper au sein de la
membrane (grâce à la fluidité membranaire) pour permettre le passage facilité de l'eau.
C'est le phénomène d'Osmose.
(cf. doc. 3, doc.B)
Transport du glucose:
Tansport actif secondaire car le gradient de concentration n'est pas favorable de la lumière du tube digestif vers la cellule. On a [Glu]extrac << [Glu]intrac.
Mécanisme:
- 1. Etat initial du transporteur (au repos): ouvert du côté extracellulaire, présente un site
spécifique au Na+, absence de site spécifique au glucose car masqué.
La fixation du Na+ induit une libération du site spécifique du glucose au sein du transporteur.
2. Fixation du glucose de manière spécifique.
Il y a alors changement de conformation du transporteur ce qui induit une ouverture de celui-ci côté intracellulaire.
3. Ouverture du transporteur côté intracellulaire.
4. L'élimination du Na+ induit le relarguage du glucose côté intracellulaire. Cela s'appel "l'effet coopératif".
5. Comme le glucose est relargué, le site spécifique de fixation de glucose disparaît, on retourne alors à l'état initial.
Maintient du potentiel membranaire:
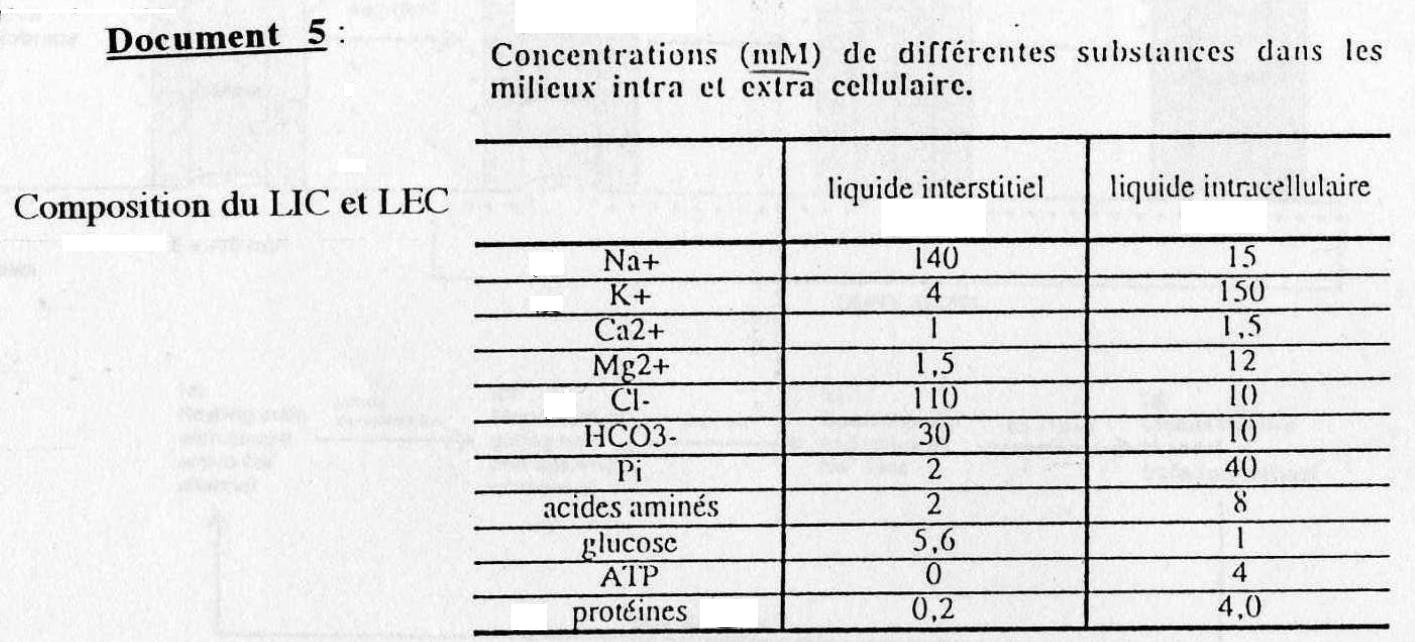
(cf. Doc. 5-6)On observe sur le doc. 5 la différente composition entre le Liquide ExtraCellulaire (L.E.C.) et le Liquide IntraCellulaire (L.I.C.).
L.E.C. = forte concentration en Na+ et Cl-
L.I.C. = forte concentration en K+ et protéines-
On mesure donc une différence de potentiel entre les 2 côtés de la membrane = "d.d.p."
Potentiel de repos et Potentiel de membrane.
Ce d.d.p. varie entre -90mV et -70mV, cela dépend de l'activité biologique de la cellule. Le potentiel de repos est essentiel à la vie de la cellule (sinon mort de la cellule).
D'autre part on constate que le Na+ a tendance à rentrer dans le milieu intracellulaire, car Ps très élevée.
Pour maintenir le potentiel de repos, la cellule doit mettre en place un système qui régule les différentes concentrations cellulaires ( comme les pompes ATPases Na+/K+).
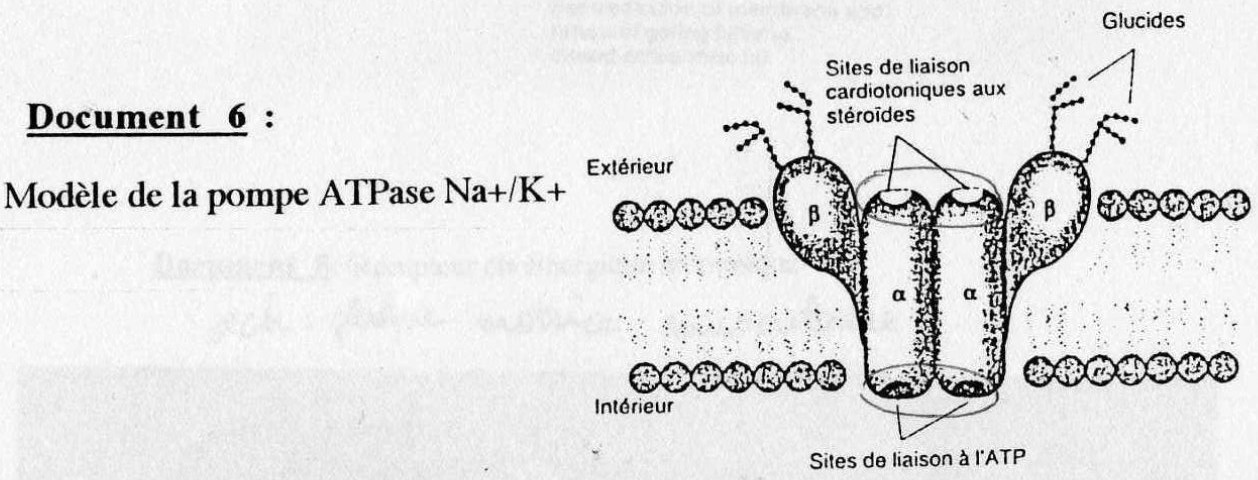
On observe sur le doc. 6 des protéines quaternaires α2 et ß2 . La sous-unité α permet l'activité ATPasique. Les récepteurs extracellulaires des sous-unités α expliquent que ces pompes
peuvent-être régulées par des hormones.
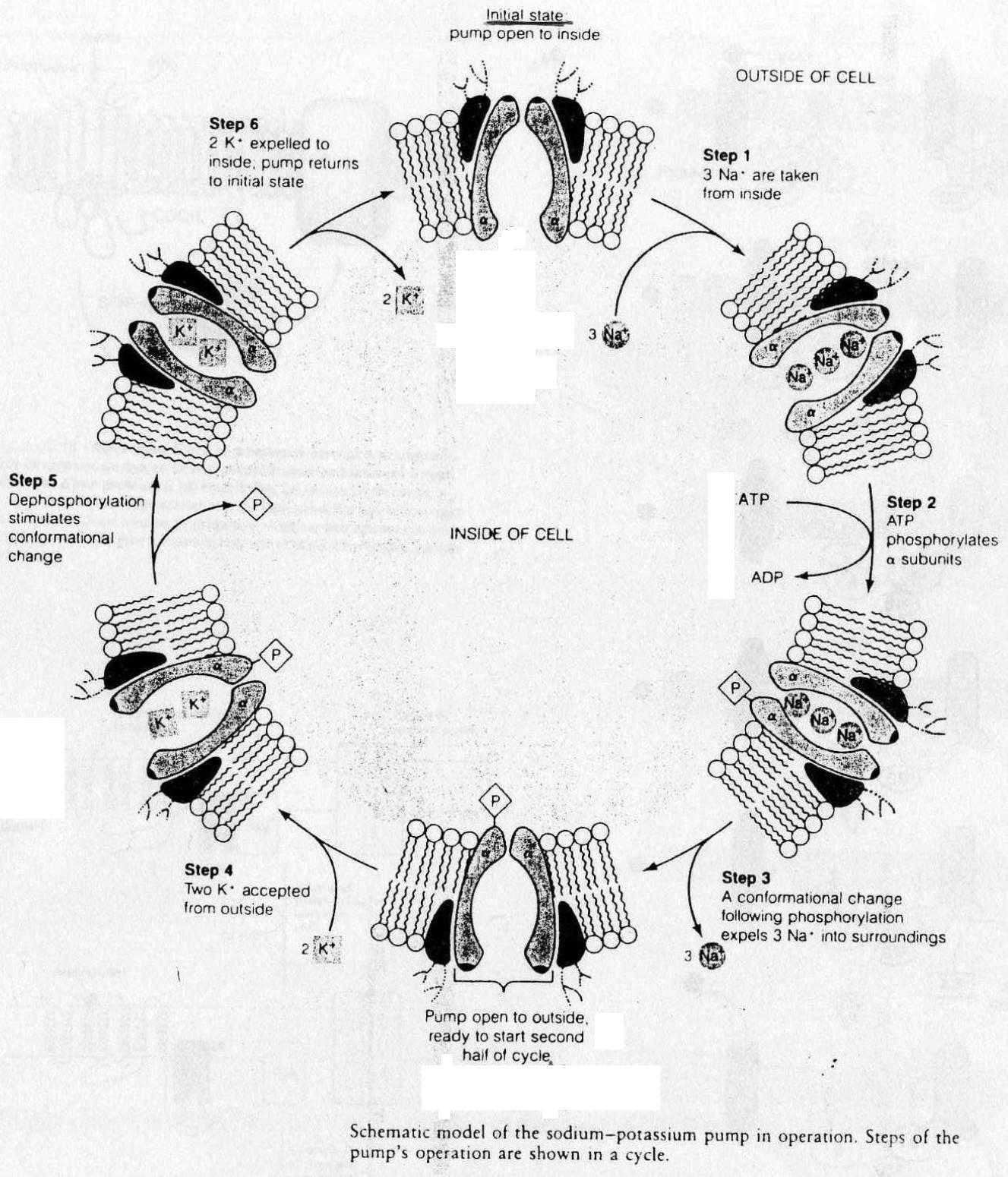
Mécanisme: (cf. Doc. 6 Bis)
A l'état de repos le transporteur est ouvert coté intracellulaire.
- 1. 3 Na+ sont fixés spécifiquement sur les sous-unités a du transporteur, coté intracellulaire.
2. Quand le site spécifique au Na+ est saturé il y a alors changement de conformation du transporteur = phosphorilation de la sous-unité a; nécessite une dépense d'énergie.
3. Il y a alors ouverture du complexe coté extracellulaire et relarguage du Na+. Apparait alors le site spécifique de fixation au K+.
4. Le transporteur prend alors en charge 2 K+.
5. Déphosphorilation de la sous-unité α, retour à la configuration initiale avec ralarguage des K+ car disparition des sites spécifiques au K+.
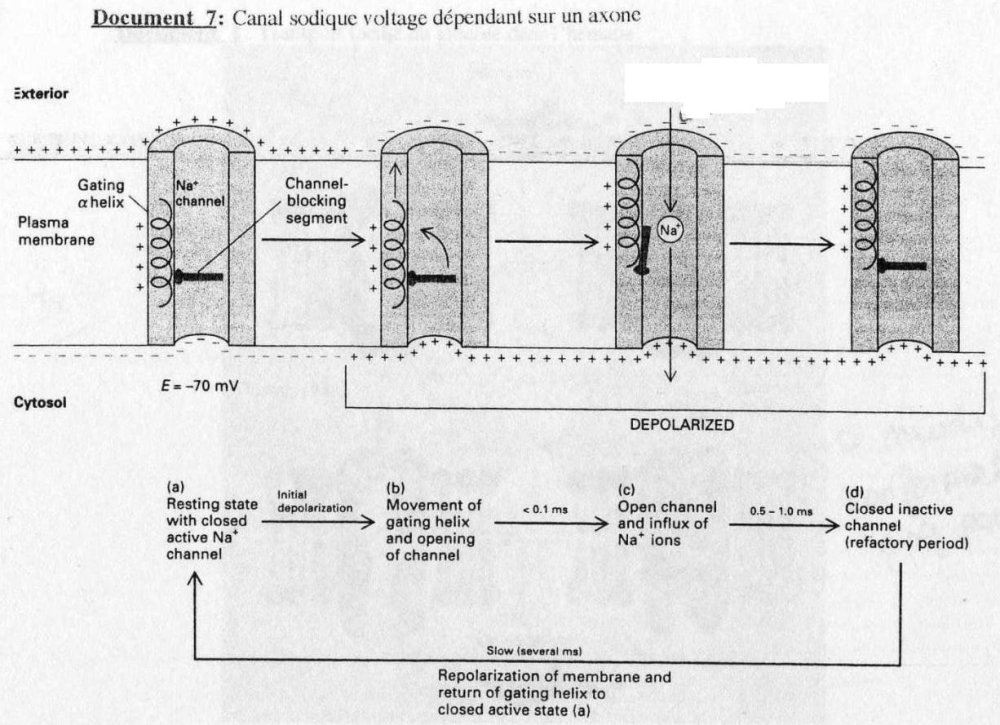
Protéine "canal" voltage dépendante:
On parle ici des axones notamment.
Ces canaux interviennent dans la conduction du message nerveux. La conduction se fait par un courant de dépolarisation locaux, entretenus par des canaux Na+ voltage dépendant. Ces
canaux peuvent-êtr sous 3 états: actif fermés, actif ouvert, inactif fermés.
Cette oscillation traduit pourquoi le potentiel d'action va du corps cellulaire à la terminaison
synaptique. Le potentiel d'action est activé que si on atteint un "seuil d'activation".
(cf. Doc. 7)
A l'arrivée du potentiel d'action, il y a une dépolarisation membranaire (potentiel d'action >
seuil d'activation). IL y a alors inversion de la charge de la membrane de l'axone. La protéine
cargée + va du coté - = ouverture du canal. On note un flux massif de Na+ ce qui se traduit
par une repolarisation locale et donc la conduction de message nerveux.
Quand le potentiel d'action a atteint le seuil maximal critique, le canal se referme = inactif. La
présence d'un état fermé inactif explique le sens de la conduction de l'influx nerveux.
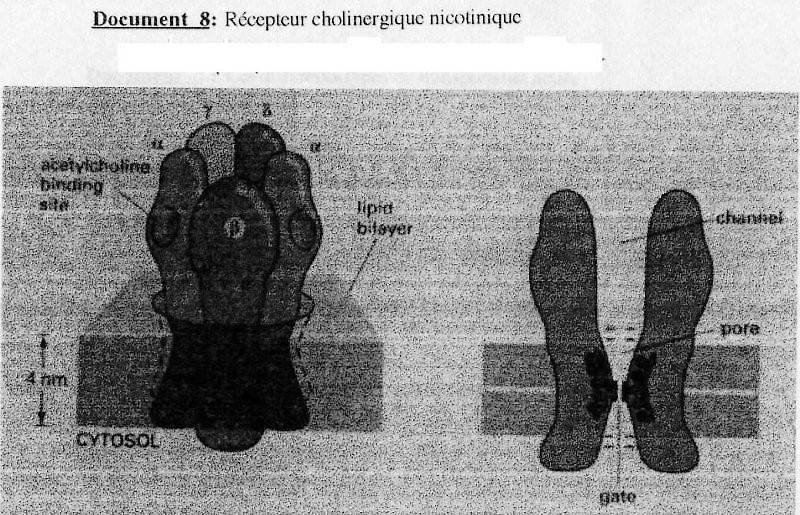
(cf. Doc. 8)
Muscle strié squelettique.
Le récepteur, l'achétylcholine (ach) se trouve sur la face post-synaptique de la cellule
musculaire. Ce canal fait partit des "récepteur canal ionotrope".
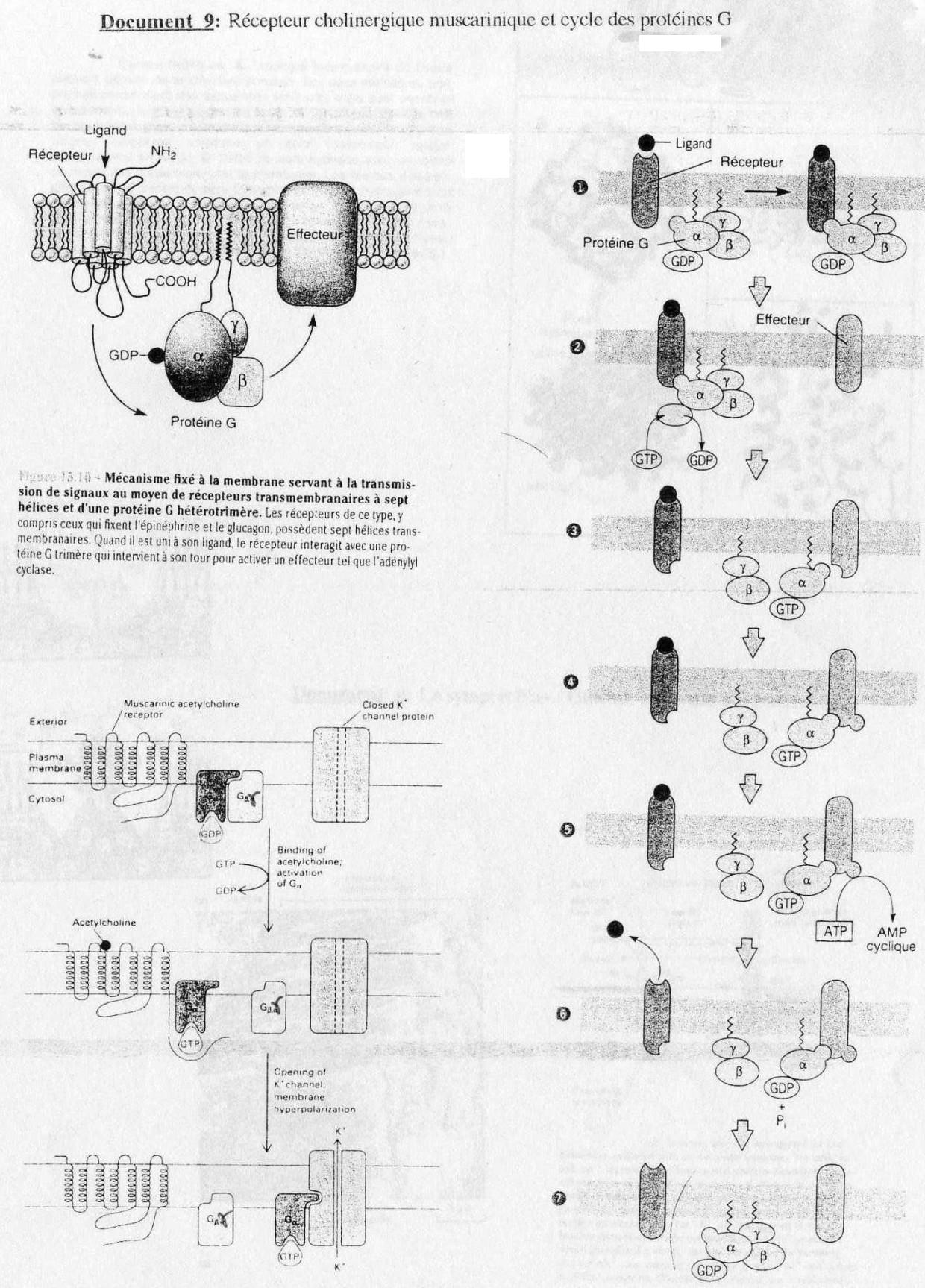
(cf. Doc. 9)
Le récepteur cholinergique est couplé à un canal K+.
La cellule s'est pourvue d'un système très complet de transport de molécules.
Ces transports impliquent majoritairement des protéines trnasmembranaires plus ou
moins spécifiques. Transporteurs, canaux et pompes pour le transport actif primaire,
qui peuvent êtr aussi impliqués dans les transports actifs secondaires ou transport facilité.
Quelques molécules diffusent librement de part et d'autre de la membrane, leur diamètre < limite d'exclusion de la membrane.
Tous ces transports sont coordonnés chez les métazoaires pour permettre un fonctionnement harmonieux des organes.
4. Annexes.
Schémas