Biologie Moléculaire et Cellulaire
LES VARIATIONS GENETIQUES
Génome =
doit-être conservé et maintenu constant (survie de l'espèce)
doit-être variable (pour s'adapter aux variations environnementales)
Ces variations du matériel génétique (ADN ou ARN) sont appelées mutations. Ces modifications
du génome:
- sont transmises directement à la descendance si reproduction asexuée.
- ne sont transmises à la descendance que si elles affectent les gamètes dans le cas d'une
reproduction sexuée.
1. Les mutations.
Mise en évidence et caractéristiques des mutations.
Mise en évidence de mutations.
- Expérience de Beadle et Tatum (1940) Notion de gènes.
Matériel: mutants de Neurospora crassa (champignon filamenteux) incapables de synthétiser l'arginine [Arg].
Hypothèse de Beadle et Tatum:
Un composé comme l'arginine doit être synthétisé par une succession d'étapes catalysées
par des enzymes différentes. On peut définir l'ordre des différentes étapes par une analyse
génétique. Pour chaque mutant on réalise un test d'auxotrophie/ prototrophie en ajoutant au
milieu de culture un des intermédiaires métaboliques connus. Deux étaient connus
concernant l'anabolisme de l'arginine: l'ornithine et la citrulline.
Analyse des mutants:

A --X--> ornithine ----> citrulline ----> Arginine
B ----> ornithine --X--> citrulline ----> Arginine
C ----> ornithine ----> citrulline --X--> Arginine
X : gène modifié
Donc a un gène = une enzyme.
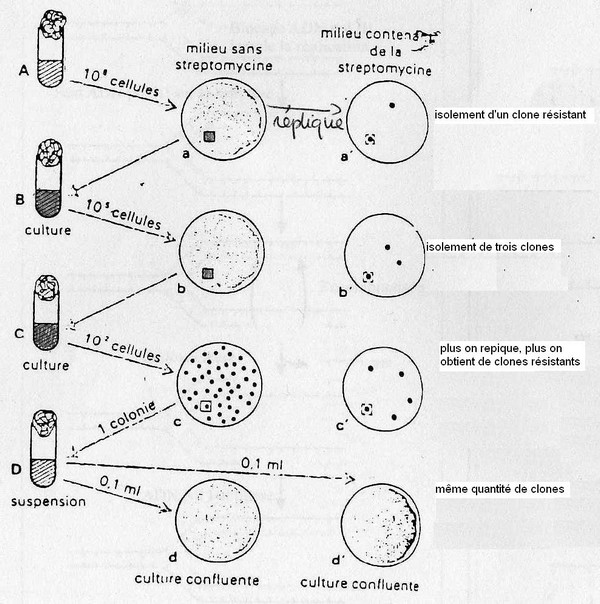
- Expérience de Lederberg.
Il voulait déterminer l'origine des mutations.
Au fur et à mesure des repiquages, le nombre de mutants augmente malgrès le fait que l'on
récupère les clones sur des populations qui sont soumises à des pressions de sélections.
Donc dans la suspension initiale il y avait des mutants => donc l' apparation des mutations est
spontanée.
Caractéristiques des mutations.
Apparition spontanée
Fréquence: 10^2-10^20 en moyenne 10^8 = fréquence d'apparition des mutations ponctuelles.
Stabilité : la mutation sera stabilisée si l'environnement apporte un avantage à la souche mutée
(cad si l'environnement maintient la pression de sélection).Sinon la mutation disparaît par
réversion (la fréquence de réversion est la même que celle d'apparition).
Généralement les mutations sont indépendantes les unes des autres.
Différents types de mutations.
Certaines altérations du génome peuvent être très importantes en taille et décelables par un caryotype altéré ou affecter des petits fragments d'ADN chromosomique ou mitochondrial dans ce cas, le caryotype est normal.
Altérations chromosomiques.
(visible chez Euvaryotes uniquement)On observe 2 types d'anomalies: en nombre ou en structure.
- Anomalies de nombre
Ces anomalies résultent le plus souvent d'une mauvaise ségrégation des chromosomes lors de la méiose de gamétogénèse.
- Anomalies de structure
Lors d'un caryotype, on récupère les chromosomes métaphasiques, on observe la taille du bras court et du bras long.

Modifications subtiles de l'ADN: mutations de l'ADN
Ces modifications ne sont pas perceptibles lors de réalisation de caryotypes. Il peut s'agir soit
d'un remplacement d'un nucléotide par un autre: mutation ponctuelle.
–transition: une base purique est remplacée par une base purique, idem pour pyrimidiques.
–transversion: une base purique est remplacée par une base pyrimidique ou inversement.
–De l'élimination de plusieurs nucléotides: délétion ou microdélétion.
–De l'addition d'un ou plusieurs nucléotides: insertion.
Conséquences des mutations.
Cas des altérations chromosomiques.
Ces altérations sont en général responsables de maladies génétiques graves chez l'homme.
Trisomie: 3 exemplaires d'un chromosome au lieu de 2 ( Syndrome de Down ou trisomie 21 = 3
chromosomes 21).
Monosomie: 1 exemplaires d'un chromosome au lieu de 2. La plupart des monosomies sont
létales sauf la monosomie X ( ex : Syndrome de Turner: individus de sexe féminin de caryotype
44 autosomes + X ).
Polyploidie: Triploïdie ( 3n chromosomes) et tétraploïdie (4n chromosomes) sont létales.
Cas des mutations plus subtiles.
Ces mutations de l'ADN affectent le plus souvent des séquences non codantes, ou des
séquences introniques ou des séquences codantes sans altérer leur fonctionnement: mutations
silencieuses. Cependant certaines altèrent le fonctionnement de séquences codantes de l'ADN:
les gènes. Dans ce cas on définit 2 types de mutations:
- les mutations conduisant à l'expression d'une protéine anormale: mutation faux-sens,
- les mutations empêchant l'expression de la protéine: mutations non sens.
Rq: certaines mutations peuvent être conditionnelles: ne s'expriment que dans des conditions
particulières exp mutants thermosensibles.
Exemples:

2. Origine des mutations.
Naturelle: due au fonctionnement même de la cellule.
Déroulement anormal de la mitose ou de la méiose.
- Erreur de ségrégation des chromosomes à la méiose/mitose,
- Résolutions altérées des chiasmas à la méiose.
Erreurs de l'ADN polymérase.
ADN polymérase incorpore 1 à 1 les nucléotides complémentaires au brin matrice. Lors de
l'addition des nt, elle commet des erreurs avec une fréquence de 10^4.
Selon la séquence, elle peut hésiter, tituber, balbutier, se tromper de nucléotide, en ajouter
quelques uns ou ne pas en insérer quelques uns.
Fidélité du processus de réplication:
- polymérisation 10^4
- activité de relecture: proof-reading: le dernier nucléotide incorporé est vérifié 10^7
- système de réparation: importance de la méthylation pour repérer le brin correct 10^10
proof-reading = fragment de Klenow (ADNpol1 + actitivté 3'-5' exonucléasique)
Donc certaines modifications ne sont pas corrigées.
Eléments génétiques mobiles.
Prophage / Virus.
Certains ADN viraux peuvent / doivent s'insérer dans l'ADN de la cellule pour que s'effectue le
cycle viral. Cette intégration peut conduire à une mutagénèse par insertion.
Conséquences: inactivation d'un gène ou surexpression du gène. Problème lorsque c'est un
gène normalement réprimé.
Mutaqènes artificiels.
Mutagènes chimiques.
- Agents modifiants les bases.
a/ Acide nitreux (HNO2) : désamination oxydative: (-NH2) → (-OH) =>Transitions
Adénine → Hypoxanthine => AT → (H)GC
Cytosine → Uraciles => CG → (U)AT
b/ Agents alkylants
Provoquent des méthylations de bases => substitutions
ex sulfonate d'éthylmethane
- Analogues de nucléotides.
ex 5 bromo-uracile existe sous 2 formes, une forme
- cétone analogue de T qui peut s'apparier avec A
- énol (plus rare) qui peut s'apparier avec G
ex 2 aminopurine
- analogue de A, s'apparie avec T
- peut former 1 seule liaison H avec C
Mutation par substitution de bases de type transition.
- Agents intercalants.
Molécules aromatiques planes peuvent s'intercaler entre les plateaux des paires de bases
adjacents ce qui conduit à l'insertion ou la délétion d'un nt ou plus.
Ex: BET ou acridine orange.
Agents mutagènes physiques.
Les radiations X et gamma sont de puissants agents mutagènes mais les lésions causées ne
sont pas exactement connues.
Les radiations UV peuvent former des dimères de pyrimidine (T-T ou C-T). Ces dimères n'ont
aucun sens pour les ADN ou ARN polymérase. Au cours de la réplication, l'ADN pol saute la
lésion ou reste bloquée.
Mais ces altérations fréquentes sont réparées par un système de réparation de l'ADN par
excision de nucléotides.
Le génome des êtres vivant évolue par transfère de matériel génétique au travers de
différentes espèces par modifications des arrangements des fragments d'ADN dans un même
génome.
3. Mécanismes des transfert d'ADN.
La transformation bactérienne (Griffith puis Averv et coll.)
La transformation bactérienne (Griffith puis Averv et coll.)
Le phénomène a été découvert par Griffith en 1928 mais n'a été compris qu'en 1944 à la suite
des expériences d'Avery, Mac Leod et Mac Carthy.
Sous forme encapsulés (Smooth), les pneumocoques présentent un fort pouvoir virulent et
conduisent à la mort par septicémie des souris infectées en 24-48 heures. Les formes sans
capsule (Rough) n'ont pas de pouvoir pathogène.
De plus, les germes isolés de la dernière souris s'avèrent être des pneumocoques S. Donc les
bactéries R ont été transformées en bactéries S grâce à un pouvoir transformant.
Avery et coll (1944) montrent que le pouvoir transformant n'est autre que l'ADN.
- les caractéristiques physiques et chimiques du pouvoir transformant correspondent à celles de l'ADN
- le pouvoir transformant est insensible à la RNAse ( nucléase hydrolysant l'ARN), mais
disparaît après traitement à la DNAse ( nucléase hydrolysant l'ADN)
Application en biologie moléculaire:
– la transformation bactérienne
– la transvection chez les Eucaryotes
Conjugaison Bactérienne.
Cas de la transmission du facteur F.
Caractéristiques mises en évidences expérimentalement: exigence d'un contact direct entre les
bactéries F+ et F-, insensible à l'action de la DNAse, seule la bactérie F- reçoit de l'information
génétique.
F+ → F-, les bactéries receveuses deviennent F+. Freq 10^6.
Donc: Conjugaison transfert unidirectionnel d'une copie d'ADN d'une bactérie donneuse vers
une bactérie receveuse (donc la bactérie donneuse conserve l'information transférée).
Nécessite le contact entre deux bactéries qui s'effectue grâce aux pilis sexuels.
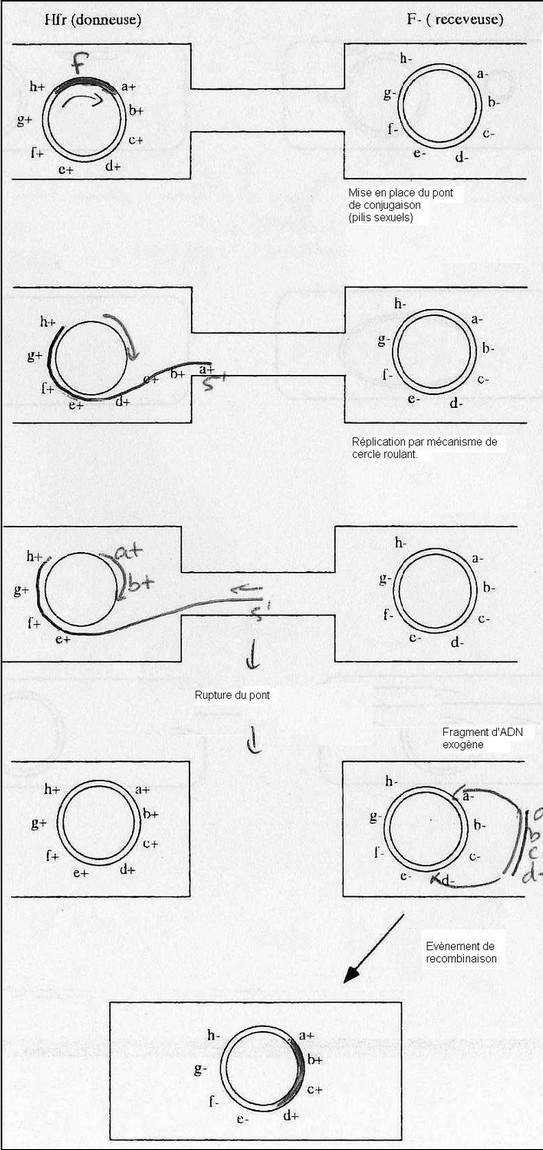
Cas de la transmission Hfr.
Obsevation d'apparition de bactéries recombinées avec une fréquence 10^1 à 10^3, acquisition
de caractères supplémentaires par transmission par le facteur F. Transmission Hfr: High Frequency of recombination.
♦ Expérience de transfert interrompu
Souche Hfr ( F+) [a+, b+, c+, d+, e+] x souche F- [ a-, b-, c-, d-, e- ]
→ Conjugaison interrompue à des temps variables par agitation

Tous les caractères chromosomiques de la bactérie donneuse peuvent être transférés.
♦ Souches F+ et Hfr
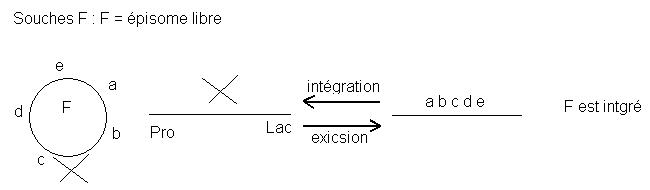
Il y a plusieurs sites d'insertion du facteur F dans le génome d'E. coll.
♦ Mécanisme du transfert Hfr
Coupure dans l'ADN du facteur F, l'extrémité 5' s'engage dans le pont de conjugaison
emportant la tête de F et le chromosome bactérien. Parallèlement, il y a néosynthèse du
chromosome bactérien ( extrémité 3' du facteur F servant d'amorce) par l'ADN polymérase
bactérienne. Le brin d'ADN transféré sert de matrice pour la synthèse d'un brin complémentaire
(exogénote). Par recombinaison avec l'endogénote, il y a acquisition des caractères a+, b+, c+,
d+ par la bactérie receveuse.
Mécanisme exclusivement bactérien.
Cette conjugaison interrompue sert à faire des cartes chromosomiques bactériennes. On
localise les gènes par rapport aux autres.
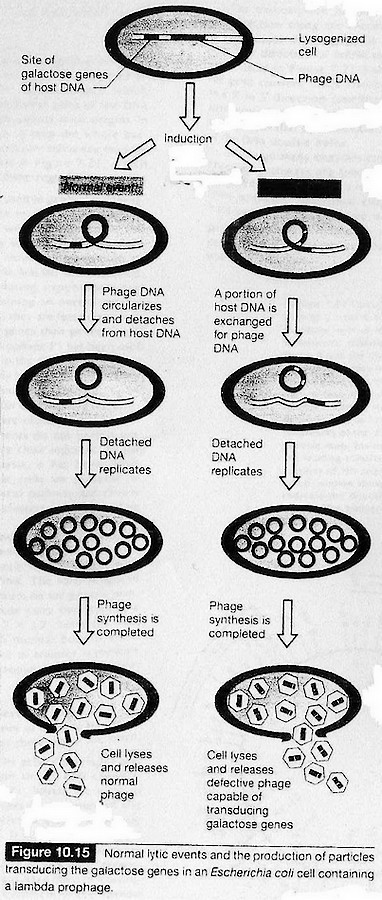
Transduction.
Découverte historique : expérience de Lederberg et Zinder (1952)

Phages transducteurs tempérés.
Découverte historique : expérience de Lederberg et Zinder (1952)
Phages transducteurs tempérés.
Cf. cours BCM
Virus transducteurs.
Exemple RSV ( Rous sarcoma virus), rétrovirus transducteur:
Au niveau des cellules animales: système analogues de transduction par une famille de virus, les rétrovirus.
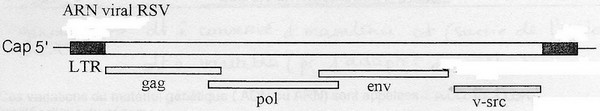
Responsables de cancers chez le poulet: notion de virus oncogènes.
Comment un rétrovirus devient oncogène:
• Rappel du cycle viral d'un rétrovirus
• Utilisation du 2ème LTR comme promoteur pour générer le 2ème ARN viral encapsidé.
Puis remaniement entre provirus.
Applications.
Utilisation des bactériophages comme vecteurs (phagemides) pour la production de simple
brins d'ADN (séquences).
Utilisation des virus comme vecteurs pour la transfection de cellules Eucaryotes, en fonction de
la multiplicité d'infection (nb de particules virales infectantes /cellule) on peut obtenir des
efficacités de 100%. De plus l'utilisation de rétrovirus permet d'obtenir directement des clones
stables.
Transposition.
Les transposons sont des éléments génétiques mobiles capables de s'intégrer dans un ADN
puis de s'exciser pour aller s'intégrer ailleurs. La transposition est permise par une activité
enzymatique la transposase codée par le transposon lui-même.
On distingue 3 types de transposons :
1) Transposons bougeant directement d'une place à l'autre (classe II)
Transposons simples
- séquences d'insertion: 2 répétitions terminales inversées encadrant le gène de la transposase.
(ex IS1, IS2, IS4, ISS pour E.coli)
- transposons complexes: de 3 types
2 répétitions terminales inversées encadrant le gène de la transposase + autres gènes
conférant au transposon des fonctions supplémentaires.
exp structure de Tn3 ( E. coli )
ex Tn5 résistance à la Kanamycine
2) MITE (Miniature Inverted-repeats Transposable Eléments)
5' GGCCAGTCACAATGG ... ~ 400 nt ... CCATTGTGACTGGCC 3'
3' CCGGTCAGTGTTACC ... ~ 400 nt ... GGTAACACTGACCGG 5'
Ces séquences sont trop petites pour coder une protéine on, pense que la transposase est
apportée par un autre transposon voisin. Ces séquences représentent 6% du génome du riz
( soit 100 000 copies) et sont présents aussi chez l'homme.
3) Rétrotransposons (Classe I)
Il transcrivent d'abord l'ADN en ARN et utilisent une réverse transcriptase pour fabriquer une
copie ADN qui s'intégrera à un autre endroit.
2 LTR encadrant une réverse transcriptase ( organisation proche des rétrovirus)
( ex Ty del levure, Bs1 du maïs...)
Mécanismes de transposition
Les 2 séquences IR du transposon s'accolent et de placent devant la séquence d'ADN cible
(certains s'insèrent au hasard).
4. Annexe.
Version Pdf
Version imprimable en pdfShémas