Microbiologie
Les bases de la Taxonomie Bactérienne
La taxonomie comprend trois domaines différents. D'une pat la classification qui consiste à regrouper les
organismes au sein d'ensembles en fonction de leurs similitudes: ensembles = « taxons ».
Ensuite la « nomenclature » qui consiste à attribuer un nom à chaque taxon.
Enfin la démarche d'identification qui utilise les 2 domaines précédents afin de reconnaître et donner un nom.
Grâce à la biochimie on a pu classer le monde des microorganismes, le génome permet d'aller encore plus loin.
1. Classifier: pourquoi et comment?
Intérêts et Structure des Classifications
Il y a trois intérêts majeurs pour classer les microorganismes:
– le classement organise une « banque de données » sur les microorganismes,
– indispensable pour identifier un nouvel isolement,
– donne accès à la phylogénie (cad le liens de parenté entre les différents organismes).
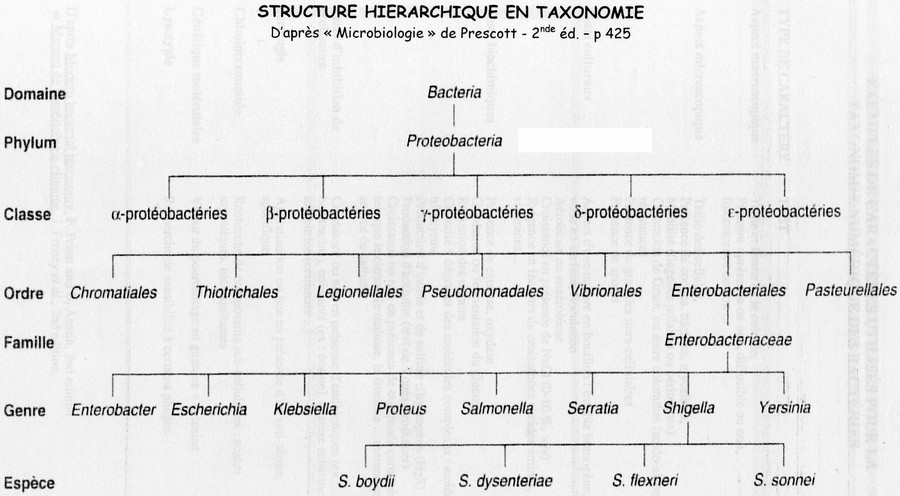
Il existe plusieurs hiérarchie au sien d'une classification, ici classification phylogénétique.
D'abord il y a les « Domaines » (Bactéries, Archéabactéries, Eucaryotes).
(...)
Différents types de classification
Il existe deux grands types: artificielle ou naturelle.
Classification artificielle
Elle est basée sur la prévalence de quelques caractères choisis arbitrairement. Ce type de classification
vient du fait que les bactériologistes n'ont pas tous la même approche de la microbiologie (médicale,
agroalimentaire, biotech, fondamental, ...). chaque domaine est tenté d'utiliser ses propres critères pour
faire de la classification ce qui implique que certaines bactéries peuvent avoir différents noms.
Ex: Erwinia hubicola (saprophyte des plantes) = Enterobacter agglomerans (bactérie intestinale)
ou encore Bacillus cereus = Bacillus thurengiensis.
Classification naturelle
On classe ici les microorganismes avec un maximum de critères sans les hiérarchiser les uns par
rapport aux autres.
Il existe deux approches différentes:
- Classification numérique phénotypique,
- Classification phylogénétique.
2. Les méthodes de la taxonomie numérique.
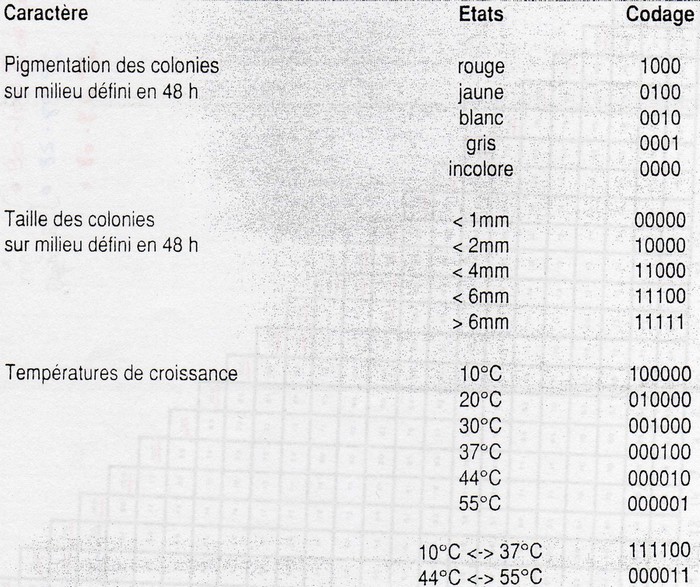
Choix des tests
Cela entraine un codage des résultats.
Ce système de codage donne des problèmes d'importance (utilisation de plus ou moins de chiffres). Or
l'implication génétique n'est pas fortement liée à l'importance des ces caractères.
Par fois quelques gènes donnent un pigment alors que cela correspond à 4 chiffres différents. Et des
centaines de gènes donnent des spores et ce critère ne correspond qu'à un chiffre. Ce n'est donc pas
équitable.
Traitements des Résultats
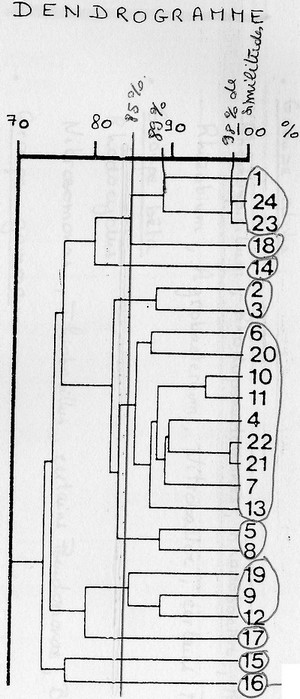
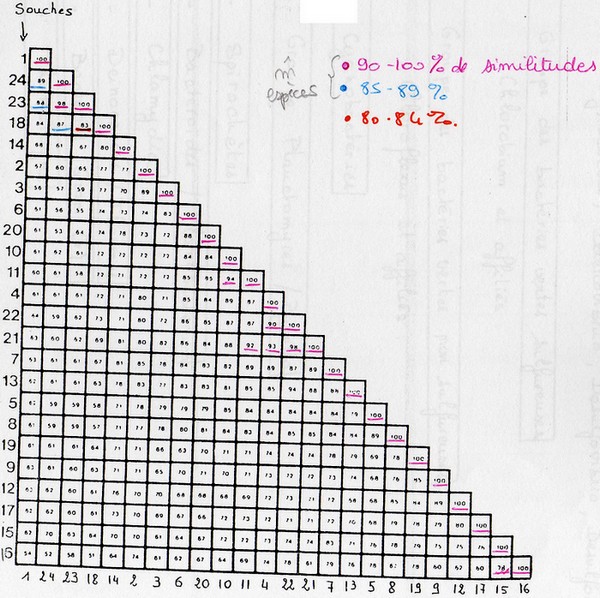
On calcul des indices de ressemblances entre chaque microorganismes de la base de données. Il y a
deux indices différents:
Coefficient de Similitudes:
nbr de caractères communs / nbr de caractères étudiés
Coefficient de Jaquard:
nbr de caractères (+) communs / nbr de caractères étudiés – caractères (-) communs
Car un caractère (-) n'est jamais certains, alors qu'un caractère (+) est sur. Le caractère peut être (-) car
souche dans de mauvaises conditions.
On considère que des souches appartiennent à la même espèce à partir de 85% de similitudes ( = limite
de l'espèce). Pour appartenir au même genre, il faut 65% de similitudes.
3. Apports de la biologie moléculaire à la classification phylogénétique .
Étude de la teneur en (G+C) de l'ADN
Justification
Si les microorganismes sont proches les uns les autres alors ils ont une teneur en GC proche.
Chez les vertébrés ce n'est pas possible car la teneur en GC varie de 35 à 45%. Or chez les bactéries
cela est intéressant car cette variation se fait de l'ordre de 25 à 75%.
Technique
(G+C)% = 2,44 Tm – 169
Tm = température à laquelle 50% de l'ADN est dénaturé.
Courbe de dénaturation thermique de l'ADN:
On étudie ici l'effet hyperchrome de l'ADN (ce qui donne le Tm et donc le %GC).
D'autre part on peut aussi faire une centrifugation, sédimentation en gradient de chlorure de Césium.
Plus le %GC est important, plus la migration se fait loin (cad plus ça tombe au fond du tube).
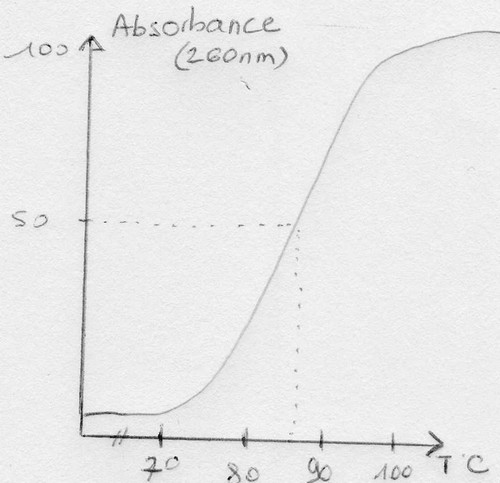
Résultats
Il faut moins de 3% d'écart pour que deux souches appartiennent à la même espèce. Il faut moins de
10% d'écart pour que deux souches appartiennent au même genre.
Attention : condition nécessaire mais pas suffisante!
Ex: spirxtaeto --> 62% de GC, pseudomonas --> 62% de GC également, mais ne sont pas de la même
espèce car physiologiquement, biologiquement et morphologiquement très différentes.
Ainsi on ne compare le %GC que pour des souches ayant le même phénotype. Cela permet de revoir la
classification.
Hybridation entre acides nucléiques
Justification
A travers le pourcentage de réassociation entre 2 ADN monocaténaires provenant de 2 souches
différentes, on peut voir le pourcentage de ressemblance entre les deux séquences.
On étudie le pourcentage d'homologie (d'hybridation), on étudie aussi le Δtm = (le TM de l'ADN hybride
= « hétéroloque ») - ( Tm de l'ADN souche référence = « homoloque »).
Technique
On travail avec l'ADN des deux souches, l'ADN de référence étant marqué par radioactivité. On mélange
les deux types d'ADN en situant le Tm à Tm – 15°C ou Tm – 30°C de la souche référence. Le mélange
est alors passer sur une colonne de chromatographie pour séparer les ADN monocaténaires des
doubles brins; les monocaténaires étant chargés et donc liés avec la colonne.
Sinon on utilise des nucléases qui agit sur le monocaténaire et qui dégrade ce qui n'a pas été hybridé ce
qui permet de garder que les doubles brins.
Résultats
Pour qu'il y ait appartenance à la même espèce il faut plus de 70% d'hybridation et un ΔTm < 5°C.
Séquençage des acides nucléiques
On s'intéresse d'abord aux ARN ribosomiques.
Justification
L'ARN reflète une partie de l'ADN.
On s'intéresse à l'ARN16s car il n'est pas trop gros (en taille l'ARN23s est trop gros et l'ARN5s est trop
petit) et il possède suffisamment d'informations.
L'ARN est une structure universelle qui se retrouve dans tout le monde vivant, on peut donc acquérir une
phylogénie de tous les être vivants.
L'ARN a une fonction identique chez tous les êtres vivants, ce qui implique que certaines séquences
soient conservées.
Il y a de l'ARN en grande quantité dans les cellules donc il s'agit d'un matériel biologique accessible.
Les gènes qui codent les ARN ribosomaux ne sont pas soumis à des « transferts horizontaux » (transfert
verticaux = transferts de gènes de mères à filles; a contrario transferts horizontaux = transferts de gènes
entre même génération, comme les plasmides!).
Technique
On a d'abord fait du séquençage partiel après digestion par une RNAse T1 qui coupe de façon
systématique des 3'G. Ce sont ainsi ces fragments qui ont été séquencé.
Méthode de SANGER: « méthode des didéoxynucléotides ». On dispose de brins d'ARN, une ARN
polymérase qui permet de faire une copie = matrice dans 4 tubes différents dans lesquels on rajoute des
nucléotides dd: ddATP, ddUTP, ddCTP et ddGTP.
L'incorporation par la matrice d'un nucléotide dd entraine le STOP de la polymérisation. On peut faire
alors une électrophorèse et on lit le résultat de la séquence en partant du nucléotide qui a le plus migré
jusqu'au bout du gel.

Séquençage totaux de 200 à 300 nucléotides d'un coup d'où l'ARN16s séquencé en 3 ou 4 fois!
Résultats
On possède des catalogues d'oligonucléotides dont certains sont spécifiques de certains groupes
bactériens= se sont des « séquences signatures ».
C'est notamment à cause de celles-ci qu'on a individualiser le groupe des Archébactéries (éclatement du
groupe des procaryotes) d'après les travaux de Woese.
Cela a amener à l'hypothèse endosymbiotique de la formation de la cellule Eucaryote. Ainsi on pense
que l'ancêtre de la sous division alpha des protéobactéries a pu être à l'origine des mitochondries, tandis
que les cyanobactéries seraient à l'origine des chloroplastes. Ainsi les mitochondries et les chloroplastes
possèdent du matériel génétique.
On a donc pu alors établir une nouvelle classification phylogénétique. Ici le critère retenus est la ressemblance de l'ARN!
Ainsi certains groupes éclatent avec des liens de parenté entre eux, tandis que d'autres se rapprochent
alors qu'ils étaient éloignés dans la classification bactérienne.
Conclusion :
A la base, on note deux approches de la classification qui s'opposent:
- L'approche Taxonomie Numérique Phénotypique qui s'intéresse aux caractères
visibles / étudiables.
- L'approche de la classification génétique qui s'intéresse à l'ADN et à la
phylogénétique entre les êtres vivants.
Problème: laquelle faut-il conserver?
La classification génétique est intéressante intellectuellement mais en pratique en elle est
difficile car pas adaptée. Donc l'approche phénotypique est tout à fait valable car plus abordable.
Aujourd'hui on mélange phénotype et génotype pour déterminer les « génomospecies ».
4. Annexes.
Document associé - pdf