Microbiologie
Multiplication des bactéries et des champignons
Etude de la croissance
Introduction
Chez les micro-organismes unicellulaires, la croissance correspond à la multiplication des individus.
L'étude de la croissance revient donc à évaluer l'augmentation de la population et s'effectue en
analysant la courbe de croissance d'une culture microbienne.
Après avoir étudié les différentes techniques d'étude de la croissance:
- évaluation de la biomasse (poids sec, turbidimétrie)
- évaluation du nombre de cellules (dénombrement en lame de Malassez, mise en culture
et comptage des colonies)
- évaluation de l'activité cellulaire (consommation d'un substrat, production d'un produit).
Nous étudierons une courbe de croissance en milieu non renouvelé (en «batch»), et nous
présenterons les cultures en batch alimenté et en continu.
1. Méthodes d'études de la croissance
Mesure de la biomasse
Dans une population bactérienne en croissance, la biomasse (ou quantité de matière vivante) augmente régulièrement. On peut l'évaluer de différentes façons.
Détermination du poids sec
II s'exprime en g/L de culture. C'est la méthode de référence, car la plus précise quand elle est bien maîtrisée, mais elle est délicate à mettre en oeuvre. (masse de microorganismes complètement déshydratés dans une culture).
Mesure d'une absorbance
- Opacimétrie
Les mesures d'opacimétrie se font entre 550 et 650 nm.
Avantages: Simples à mettre en oeuvre; Peu couteux.
Inconvénients:Domaine de linéarité : λ= 650nm; A linéaire jusqu'à A = 0,5 --> A réelle du milieu. Inapplicable lorsque le milieu est trouble.
Valeur estimée par excès: on compte ici les cellules vivantes et les cellules mortes donc on surestime la quantité de biomasse.
- Turbidimétrie:
La turbidité se mesure par détermination de l'épaisseur minimale de suspension empêchant la vision nette d'une mire, ou par comparaison avec un étalon.
- Néphélométrie:
Mesure du nombre de cellules
Le nombre de cellules d'un milieu peut-être évalué par des méthodes directes cad compter directement le nombre de cellules.
Comptage à l'hématimètre
Compteur de particules
C'est un système automatique qui dénombre les bactéries qui passent entre 2 électrodes au niveau d'une micropipette.
Inconvénients: Ce système est peu adapté à la taille des bactéries. De plus il dénombre les cellules vivantes
et mortes. Donc par excès.
Des marqueurs fluorescents à l'acridine (qui se fixe à l'ADN) permettent de différencier les cellules vivantes
(vertes) des cellules mortes (rouges).
On peut également évaluer le nombre de bactéries par des méthodes indirectes cad par culture de cellules.
Numérations en milieu solide
Cette technique consiste à réaliser des dilutions décimales du produit à étudier et à ensemencer soit 1 mL en profondeur d'un milieu solide en surfusion, soit 0,1 mL en
surface à l'aide d'un râteau. (2 boîtes par dilution)
Les résultats sont donnés en UFC.

Avantages des méthodes indirectes: On ne compte que des cellules vivantes.
Inconvénients des méthodes indirectes: On ne fait pas de différences entre une colonie issue d'un microorganisme ou issue d'un amas de microorganismes. On augmente le pourcentage d'erreurs. Il y a une étape d'incubation donc les résultats ne sont pas immédiats.
Détermination de l'activité
II est possible de suivre par des techniques biochimiques la consommation d'un substrat (ex : suivi de la
consommation de glucose par dosage enzymatique) ou l'apparition d'un produit (ex:dosage de l'éthanol).
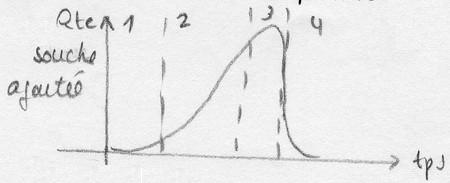
On peut également suivre l'activité des micro-organismes en étudiant le maintient d'un paramètre régulé.
Ex: le pH est en général un paramètre régulé au cours d'une fermentation. Donc sa valeur ne donne pas
d'information sur l'activité des micro-organismes, mais l'action nécessaire pour maintenir le pH constant
sera , elle, représentative du niveau d'évolution de la réaction de fermentation. Ainsi, en suivant la
quantité de base injectée par unité de temps, la courbe obtenue sera:
2. Suivi de la croissance en milieu non renouvelé
Un milieu non renouvelé est un milieu pour lequel on n'apporte pas de nouveau substrat en cours de culture ainsi la croissance est limitée et elle s'arrête par épuisement des substrats et accumulation des déchets. En pratique, c'est ce qui se produit dans un erlen ou un tube de bouillon.
La courbe de croissance
La croissance cellulaire se fait par divisions binaires successives selon le schéma:
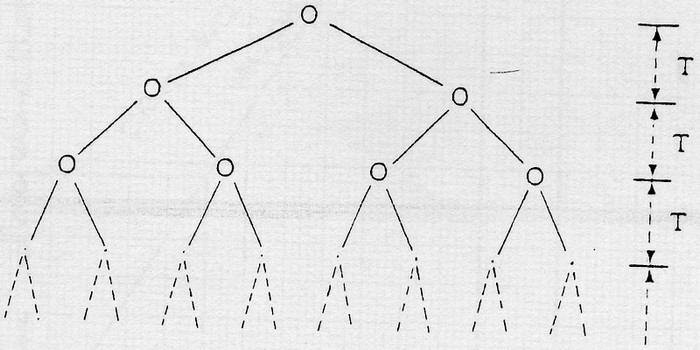
T étant le temps s'écoulant entre 2 divisions.
On peut écrire l'équation suivante:
N: nombre de cellules totales
N0: nombre de cellules totales à t=0
N = N0 x 2t/T
=> ln N = ln N0 + t/T ln 2
ln N = ln N0 + ln 2 / T x t
avec ln N0 = constante et ln 2 / T = variable car T est fonction des conditions de cultures.

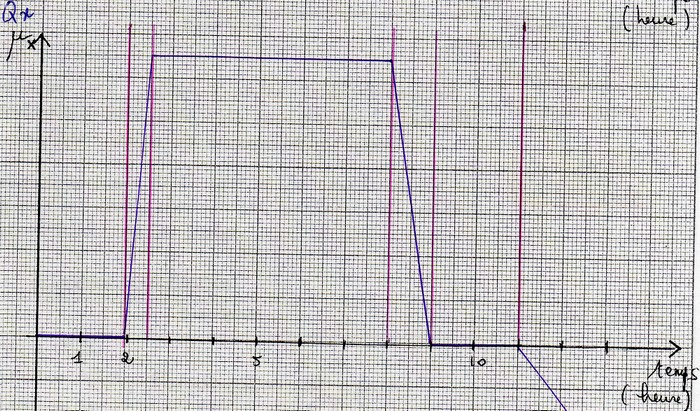
Les différentes phases de la croissance
On peut distinguer 4(+2) phases lors de la croissance d'un micro-organisme en milieu non renouvelé.
1- La phase de latence

1'- La phase d'accélération

2- La phase de croissance exponentielle

2'- La phase de décélération

3- La phase stationnaire

4 - La phase de déclin

Les paramètres de la croissance
On définit principalement 2 paramètres pour évaluer la croissance microbienne:
Vitesse Spécifique de Croissance μx = Qx
Qx = « quotient de Biomasse »
On évalue l'accroissement de la biomasse (x = nbr d'individu en g/L)
Définition: accroissement de la Biomasse (dx) / unité de temps (dt) ramené à l'unité de biomasse (x)
Qx=(dx / dt) x 1/x => Qx=(dx / x) / 1/dt => Qx=d lnx x 1/dt => d lnx=Qx x dt
=> ∫ d lnx=∫ Qx dt => [ lnx2 – lnx1]=[Qx (t2 – t1)]
=> Qx=(lnx2 – lnx1) / (t2 – t1) = coefficient directeur de lnx = f(x) à tout moment!
Qx peut se mesurer à n'importe quel moment, Le plus souvent on calcul Qx pendant la phase
exponentielle car à ce moment Qx est maximal.
Temps de Génération : G
C'est le temps de doublement de la population pendant la phase de croissance exponentielle, cad le
temps qui sépare deux divisions cellulaire (cad Tmin = G).
G = t2 – t1 quand x2 = 2x1
Qx expo = (lnx2 – lnx1) / (t2 – t1) or en phase exponentielle x2 = 2x1 d'où:
Qx expo = (ln 2x1 – lnx1) / G => Qx expo = Ln 2 + lnx1 – lnx1 / G => G = ln 2/Qx expo
Détermination graphique de G:
On prend lnx1, ce qui nous donne t1 puis pour avoir t2 on prend un second point avec ln 2 + ln x1, car il
faut x2 = 2x1 cad ln x2 = ln 2 + ln x1
Donc si on a ln x2 alors on a t2, alors G = t2 – t1
Suivi de Fermentation et calculs de rendements
Lorsque l'on réalise une production en fer menteur on peut suivre en plus de l'évolution de la biomasse,
l'utilisation du substrat et l'apparition du produit. On calcul alors des rendements de production:
Vitesse de Consommation du Substrat:
rs = d[S] / dt exprimé en qté S / t (ex: g/L / H)
Vitesse d'Apparition du Produit:
rp = d[P] / dt exprimé en qté P / t (ex: g/L / H)
Vitesse Spécifique de consommation du Substrat:
ramené à l'unité de Biomasse
Qs = ( d[S] / dt ) x ( 1/x )
Vitesse Spécifique de consommation du Substrat:
Qp = ( d[P] / dt ) x ( 1/x )
Rendement:
Y x/s = rendement en biomasse par rapport au Substrat
Y x/s = d[x] / d[S] = rx / rs exprimé en quantité de Biomasse par quantité de substrat consommé
Y p/s = d[P] / d[S]
Rmq: La production d'un métabolite peut se faire de 3 façons différentes:
Association entre la croissance et la biosynthèse: METABOLTTES PRIMAIRES
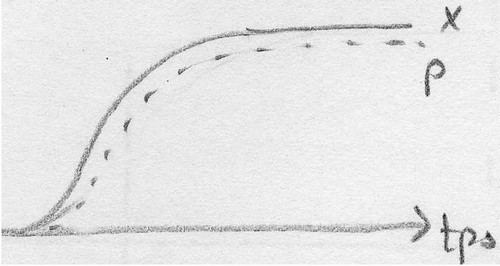
Association partielle entre la croissance et la biosynthèse

Dissociation entre croissance et biosynthèse: METABOLTTES SECONDAIRES
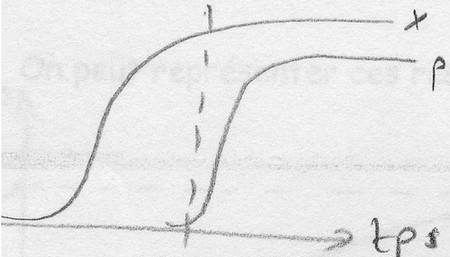
P = antibiotiques ou toxines la plus part du temps
Facteurs de variation de la croissance
La cinétique de croissance d'un micro-organisme est influencée par:
- des facteurs qualitatifs: nature de la source de C, N ...
- des facteurs quantitatifs: [S], température, pH ...
Les facteurs physico-chimiques
La teneur en O2:
Détermine le type de métabolisme (fermentaire ou respiratoire); influe sur la production des métabolites
primaires (comme l'éthanol); influe sur la vitesse de consommation du Substrat (on joue sur l'agitation du
milieu).
Le pH:
Influe sur la vitesse de catalisation des enzymes, sur la vitesse de synthèse des métabolites, influe sur
la vitesse de croissance (lors de pH acides la croissance peut-être rapidement inhibée).
La température:
Influe la dure de la phase exponentielle mais ne modifie pas la quantité de biomasse finale. Peut aussi
influencer le s métabolites synthétiser car la température peut-être un facteur de stress.

La concentration et la nature du substrat
Évolution de la courbe de croissance en fonction de [S]:
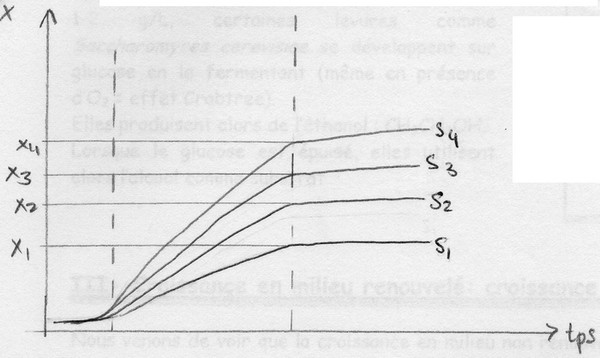
On peut représenter ces résultats en traçant : μxexpo = f ([glucose])
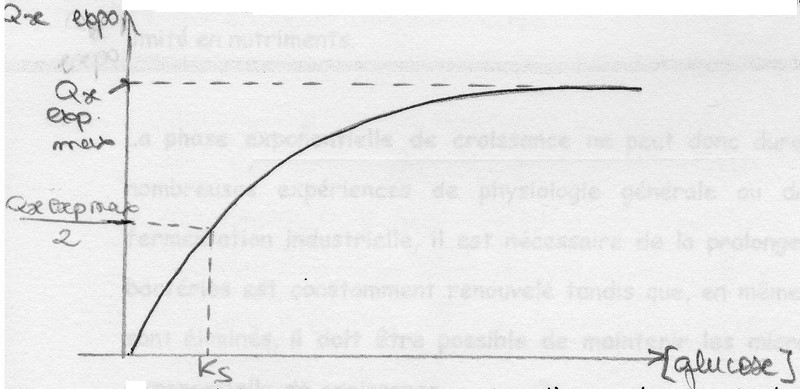
On constate que μxexpo augmente jusqu'à une valeur maximale qui est μxexpo Max.
Lorsque μxexpo = μxexpo Max, la [glucose] est telle que le glucose n'est plus le facteur limitant de la croissance.
La représentation est régie par l'équation de Monod: Qxexpo = Qxexpo max [S] / Ks + [S]
Ks = constante de saturation = traduit l'affinité du microbe pour le substrat (si Ks élevé = affinité faible et vice versa).
Évolution de la courbe de croissance en fonction de la nature de S: phénomène de diauxie
Exemple 1:
Milieu synthétique contenant:
- un ose utilisable directement (ex: glucose)
- un diholoside nécessitant une adaptation enzymatique (ex: maltose ou fructose).
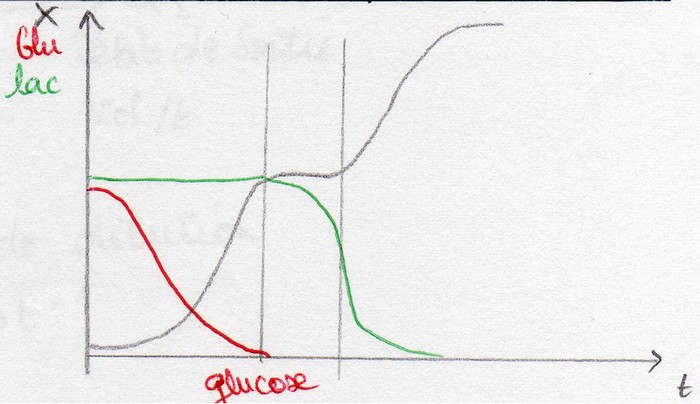
Exemple 2:
Pour une concentration en glucose supérieure à 1-2 g/L, certaines levures comme Saccharomyces cerevisiae se développent sur glucose en le fermentant (même en présence d'O2 - effet Crabtree). Elles produisent alors de l'éthanol: CH3CH2OH.
Lorsque le glucose est épuisé, elles utilisent alors l'alcool comme substrat.

3. Croissance en milieu renouvelé: croissance continue
Nous venons de voir que la croissance en milieu non renouvelé (ou croissance en discontinue) est un
système clos où le cycle de développement est le résultat d'une croissance dans un milieu limité en nutriments.
La phase exponentielle de croissance ne peut donc durer que quelques heures. Or, pour de
nombreuses expériences de physiologie générale ou dans un but plus pratique pour la fermentation
industrielle, il est nécessaire de la prolonger. Si le milieu où se multiplient les bactéries est
constamment renouvelé tandis que, en même temps, les produits du métabolisme sont éliminés, il doit
être possible de maintenir les micro organismes indéfiniment en phase exponentielle de croissance.
Définition
On appelle culture continue, un système ouvert dans lequel la population de cellules est maintenue constamment dans un milieu équilibré en nutriment et en concentration cellulaire. Pour cela une partie de la culture est retirée et remplacée par du milieu neuf.
Principe
Paramétrage
Dans un tel système où la culture reste sous un volume constant (V), tout en recevant un débit (en
vol/unité de temps) de milieu neuf, la concentration cellulaire varie en fonction de la vitesse spécifique
de croissance en phase exponentielle ( Qx max.) de l'espèce:
où X est la biomasse, D le taux de dilution (rapport du débit d'entrée sur le volume de milieu exprimé en t-1).
Trois cas possibles
On peut distinguer trois éventualités:
• Qx max < D: l'effet de dilution est supérieur à l'incidence de la croissance. Dans ces conditions, la
biomasse tend vers 0, car dX/dt <0. Au bout d'un certain temps on retrouve dans le fermenteur un
milieu pur.
• Qx max = D: le taux de dilution est calculé pour que la biomasse reste constante et maximale dans les
conditions choisies (Qx max) dX/dt = 0.
Certains dispositifs ont été conçus pour correspondre à cette situation : ce sont les TURBIDOSTATS.
• Qx max > D: l'effet de dilution est moins important que celui de la croissance. dX/dt > 0. Il va exister
donc un facteur limitant et Qx < Qx max, La biomasse tend à augmenter, elle peut être autorégulée
pour la maintenir constante. Ceci a permis de mettre au point le CHEMOSTAT.
Deux grands systèmes
Le turbidostat
Ce dispositif assure une culture continue à Qxmax et une biomasse constante, X, qui sera choisie en phase exponentielle et restera constante grâce à une régulation de D. On autorégule D en fonction de la biomasse mesurée en sortie de système.
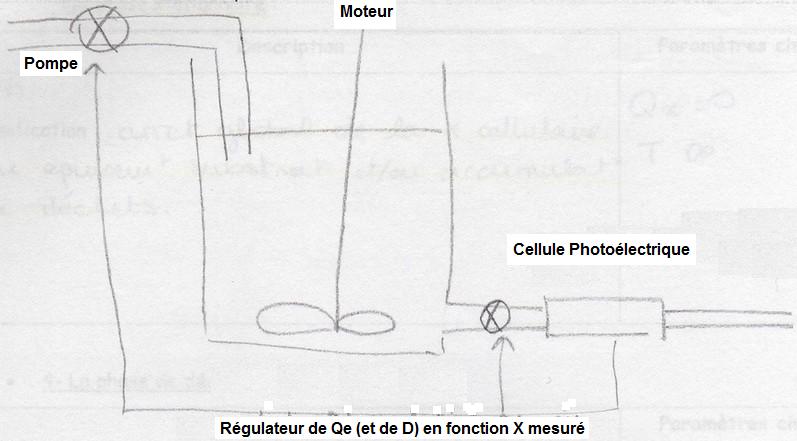
Le chémostat
Dans cet appareil, Qx n'atteint jamais Qx max. Sa valeur est liée à la concentration d'un facteur limitant dans la chambre de croissance, tous les autres nutriments étant en excès. Cette concentration dépend du débit de milieu neuf, donc de D. Il est ainsi possible de choisir Qx la plus convenable en faisant varier la valeur D.
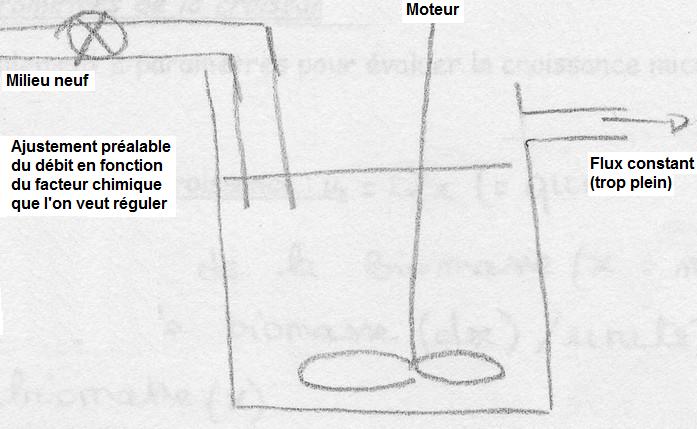
Conclusion:
Avantage de la croissance continue
vitesse de croissance contrôlée et indéfiniment maintenue ainsi que la biomasse.
Inconvénients de la croissance continue
production de métabolites secondaires délicates,
sélection de mutants en cours de fermentation
problèmes de contamination inhérents à une longue période de culture.